引用本文: 史平玲, 魏圓夢, 黃子旭, 盧聰, 楊琪翔, 李盼, 鄔成業, 宋宗明. 轉膠蛋白-2抑制高糖誘導的小膠質細胞炎癥反應的轉錄組測序分析. 中華眼底病雜志, 2023, 39(2): 153-162. doi: 10.3760/cma.j.cn511434-20210716-00381 復制
版權信息: ?四川大學華西醫院華西期刊社《中華眼底病雜志》版權所有,未經授權不得轉載、改編
視網膜小膠質細胞是視網膜內免疫監測細胞,靜止狀態下其通過吞噬和控制低度炎癥幫助維持視網膜的組織穩態。但高血糖引起的長期組織應激可導致小膠質細胞過度反應,并產生促炎細胞因子和趨化因子引起慢性炎癥[1]。在糖尿病視網膜病變(DR)中小膠質細胞活化導致炎癥增加和免疫代謝紊亂[2]。鏈脲佐菌素誘導的DR模型小鼠,小膠質細胞活化并聚集在視網膜色素上皮層,產生的白細胞介素(IL)-6進一步促進小膠質細胞的募集和腫瘤壞死因子(TNF)-α的釋放,加重血視網膜屏障破壞[3]。通過抑制小膠質細胞引發的視網膜炎癥,減少高糖誘導的細胞外調節蛋白激酶1/2-核因子(NF)-κB通路的激活,能夠減輕DR[4]。轉膠蛋白-2(TAGLN2)為肌動蛋白(actin)細胞骨架結合蛋白,屬于轉膠蛋白超家族[5];其主要定位于細胞胞漿,可通過直接調控相關基因表達而影響細胞表型[6]。TAGLN2在平滑肌細胞及成纖維細胞中高表達,參與血管生成、骨架重塑以及細胞遷移、凋亡及增生等功能調控,能夠調節T細胞激活,在B細胞中也參與了T細胞-B細胞結合物的穩定[5-8]。TAGLN2在脂多糖刺激的巨噬細胞中表達高度上調,并促進巨噬細胞的吞噬[9]。重組TAGLN2蛋白能夠增強樹突狀細胞抑制腫瘤生長和轉移的功能[10]。因此TAGLN2在淋巴細胞、巨噬細胞和樹突狀細胞的免疫反應中都發揮了重要的作用。但是TAGLN2是否在高糖誘導的小膠質細胞免疫反應中起作用及其作用機制鮮見報道。轉錄組測序分析技術為細胞功能研究提供了新的方法[11]。本研究利用慢病毒沉默和過表達BV2小膠質細胞(以下簡稱為BV2細胞)中的TAGLN2基因,通過轉錄組測序和生物信息學分析闡明TANLN2調控高糖誘導的小膠質細胞炎癥反應的分子機制,為DR的治療提供新的策略。現將結果報道如下。
1 材料和方法
1.1 主要材料
BV2細胞(國家實驗細胞資源共享服務平臺)。D-葡萄糖(Glu)、甘露醇(Man)(德國Merck公司),嘌呤霉素、蛋白裂解液、山羊血清(北京索萊寶科技有限公司),胎牛血清(美國Gibco 公司),脂質體Lipofectamine? 3000、Trizol試劑、4',6-二脒基-2-苯基吲哚(DAPI)(美國Invitrogen公司),蛋白酶抑制劑(美國MCE公司)。PrimeScript RT reagentKit with gDNA eraser試劑盒、SYBR Rremix Ex Taq II Kit試劑盒(日本TaKaRa公司),二喹啉甲酸(BCA)蛋白濃度測定試劑盒(北京索萊寶科技有限公司),Premixed Multiplex Kit Mouse Custom 8-Plex Kit試劑盒(北京曠博生物技術股份有限公司)。質粒載體:pHelper1.0、pHelper2.0(上海吉凱基因科技有限公司),psPAX2、PMD2.G(漢恒生物科技有限公司)。抗體:抗TAGLN2兔多克隆抗體、抗CD11b兔多克隆抗體(美國Abcam公司),抗Iba-1羊多克隆抗體(美國Novus公司),辣根過氧化物酶(HRP)標記羊抗兔免疫球蛋白G(IgG)、 硫氰酸熒光素(FITC)標記的羊抗兔IgG(FITC-IgG)(中國博奧森生物技術有限公司),藻紅蛋白(PE)標記的驢抗羊IgG[PE-IgG,愛必信(上海)生物科技有限公司]。儀器設備:NanoVue紫外/可見光分光光度計(美國GE Healthcare公司),Agilent 2100 bioanalyzer生物分析儀(美國賽默飛世爾科技公司),熒光顯微鏡(日本Olympus公司),BD FACSCanto流式細胞儀(美國Becton Dickinson公司)。
1.2 細胞培養和分組
培養的BV2細胞分為Man組、Glu組、過表達對照(Con)Glu組(Con Glu組)、過表達TAGLN2 Glu組(TAGLN2 Glu組)、沉默Con Glu組(shCon Glu組)、沉默TAGLN2 Glu組(shTAGLN2 Glu組)。
Man組BV2細胞置于含25 mmol/L Man、25 mmol/L D-Glu、10%胎牛血清的改良Eagle培養基(DMEM培養基)中培養; Glu組、Con Glu組、TAGLN2 Glu組、shCon Glu組、shTAGLN2 Glu組BV2細胞置于含50 mmol/LGlu和10%胎牛血清的DMEM培養基中培養。各組細胞均于37 ℃、5% CO2培養箱中孵育24 h,收集細胞。
1.3 重組慢病毒包裝
根據篩選出沉默效率最高的siRNA序列設計合成短發夾RNA(shRNA)序列,將沉默TAGLN2基因的shRNA序列CCGGGCCGTGAGAACTTCCAGAACTCGAGTTCTGGAAGTTCTCACGGCTTTTT,陰性對照shRNA序列CCGGTTCTCCGAACGTGTCACGTCTCGAGTTCTGGAAGTTCTCACGGCTTTTT分別與慢病毒載體GV248進行連接,并與輔助質粒pHelper1.0、pHelper2.0在293T細胞中包裝成慢病毒GV248-shRNA,感染BV2細胞,4 μg/μl嘌呤霉素篩選,分離單克隆細胞并擴大培養。
合成TAGLN2基因(NM_178598.2)的CDS區序列并與慢病毒載體pHBLV質粒進行連接,與輔助質粒psPAX2、PMD2.G包裝成慢病毒pHBLV-TAGLN2,pHBLV空質粒與輔助質粒包裝成對照慢病毒pHBLV-Con,分別感染BV2細胞,4 μg/μl嘌呤霉素篩選,分離單克隆細胞并擴大培養。
1.4 實時聚合酶鏈反應(RT-PCR)檢測細胞中各因子mRNA相對表達量
采用Trizol試劑提取細胞總RNA,NanoVue紫外/可見光分光光度計檢測RNA樣本濃度,PrimeScript RT Reagent Kit with gDNA Eraser試劑盒將RNA反轉錄成cDNA,使用SYBR Rremix Ex TaqⅡ Kit試劑盒和7500RT-PCR系統進行RT-PCR。每組4個樣本,每個樣本重復3次。相關引物由生工生物工程(上海)股份有限公司合成(表1)。反應條件: 95 ℃預變性30 s,循環1次;95 ℃變性5 s,60℃退火延伸40 s,循環40次;95 ℃變性15 s,60 ℃退火1 min,95 ℃變性15 s,循環1次。以β-actin作為內參照,采用 2?ΔΔCt方法進行結果分析。
1.5 蛋白免疫印跡法檢測細胞中TAGLN2蛋白相對表達量
利用組織蛋白裂解液和蛋白酶抑制劑的混合液提取BV2細胞蛋白,BCA法測定蛋白濃度,取等量蛋白進行十二烷基硫酸鈉聚丙烯酰胺凝膠電泳,并行蛋白質分離且轉印至硝酸纖維素膜上,含10%脫脂牛奶的磷酸鹽緩沖液(PBS)室溫封閉2 h,1∶200稀釋的抗TAGLN2兔多克隆抗體4℃孵育過夜。轉印膜于含0.5%吐溫-20的PBS溶液(PBST)中洗膜10 min,重復3次;1∶3 000稀釋的HRP-IgG室溫孵育2 h,PBST洗膜3次。高敏化學發光試劑盒檢測各組蛋白相對表達量。每組3個樣本,重復1次。
1.6 免疫熒光染色檢測Glu組、Man組BV2細胞中Iba-1表達
4%多聚甲醛固定細胞爬片10 min,0.2% Triton X-100通透30 min,滴加10%山羊血清于室溫下封閉30 min。加入1∶100稀釋的一抗抗CD11b兔多克隆抗體、抗Iba-1羊多克隆抗體,4℃孵育過夜,PBST洗滌3次;加入1∶200稀釋的二抗羊抗兔FITC-IgG、驢抗羊PE-IgG,DAPI避光染色30 min,防熒光淬滅封片劑封片,熒光顯微鏡觀察。細胞中CD11b呈綠色熒光,Iba-1呈紅色熒光,細胞核呈藍色熒光。每組3個樣本,1次免疫熒光染色。
1.7 AimPlex流式高通量多因子檢測BV2細胞中IL-1β、TNF-α、IL-6、干擾素(IFN)-γ、IL-17蛋白表達
收集各組細胞上清液,4 ℃條件下以離心半徑8.6 cm、12 000 r/min離心5 min,25 μl上清液和25 μl樣品稀釋液混勻制成稀釋樣品,標準品稀釋液梯度稀釋標準品。使用Premixed Multiplex Kit Mouse Custom 8-Plex Kit試劑標記標準品或稀釋樣品,依次加入45 μl/孔已包被抗體的混合微球和45 μl/孔的標準品或稀釋樣品,封板,37 ℃、700 r/min避光震蕩60 min。加入1倍洗液100 μl/孔,重復3次;加入1倍生物素標記的二抗25 μl/孔,封板,37℃、700 r/min避光震蕩30 min。加入1倍洗液100 μl/孔,重復2次。加入1倍讀數液200 μl/孔,反復吹吸實驗孔懸浮微球并轉移入5 ml流式細胞管中,使用BD FACSCanto流式細胞儀488 nm激光對反應中的不同大小、不同熒光強度的微球群進行檢測。每組3個樣本,1次檢測。
1.8 轉錄組測序分析
每組設3個生物學重復,Trizol Reagent試劑提取細胞總RNA,NanoDrop超微量分光光度計、Agilent 2100 bioanalyzer生物分析儀檢測RNA濃度、質量,質檢合格構建mRNA文庫,BGI-seq500 平臺進行轉錄組高通量測序,原始測序數據通過SOAPnuke去除低質量、接頭污染以及未知堿基N含量>5%的讀數得到高質量序列。應用HISAT(v2.0.4)、Bowtie2(v2.2.5)分別將高質量序列比對到美國國立生物技術信息中心參考基因組序列GCF_ 000001635. 26_GRCm38.p6、GCF_ 00000 1635.26,得到比對結果。計算每兩個樣品之間所有基因表達量的Pearson相關系數。采用轉錄組學測序技術篩選各組顯著差異表達基因(DEG),篩選標準為|log2[差異表達倍數(FC)]|≥1且P≤0.05[11]。通過RSEM(v1.2.12)軟件計算基因表達水平,DESeq2(v1.4.5)軟件篩選DEG。采用GraphPad Prism軟件繪制火山圖描繪顯著上調基因(log2 FC≥1,P<0.05)和下調基因(log2 FC≤-1,P<0.05)表達的分布情況;R軟件中pheatmap函數繪制聚類熱圖,分析DEG變化;R軟件中的phyper函數進行基因注釋(GO,http://www.geneontology.org/)和京都基因與基因組百科全書(KEGG,https://www.kegg.jp/)富集分析,富集顯著性標準為P<0.05。通過STRING數據庫(https://string-db.org)和Cytoscape軟件進行蛋白-蛋白互作網絡(PPI)分析,圖中節點大小代表該基因權重的高低,節點越大,基因權重越高。
1.9 統計學分析
采用GraphPad Prism軟件進行統計學分析。呈正態分布的計量資料以均數±標準差( ±s)表示。組間數據比較采用獨立樣本t檢驗。P<0.05為差異有統計學意義。
±s)表示。組間數據比較采用獨立樣本t檢驗。P<0.05為差異有統計學意義。
2 結果
2.1 高糖誘導小膠質細胞模型建立
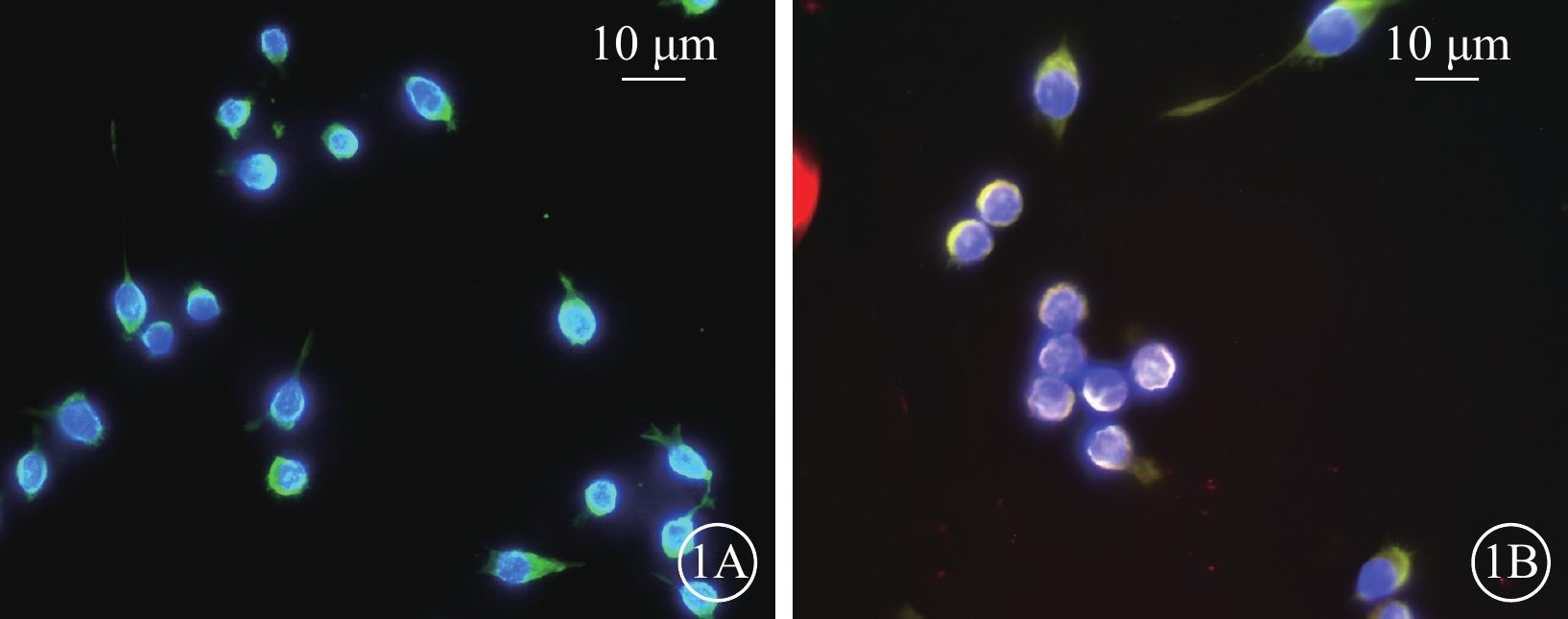
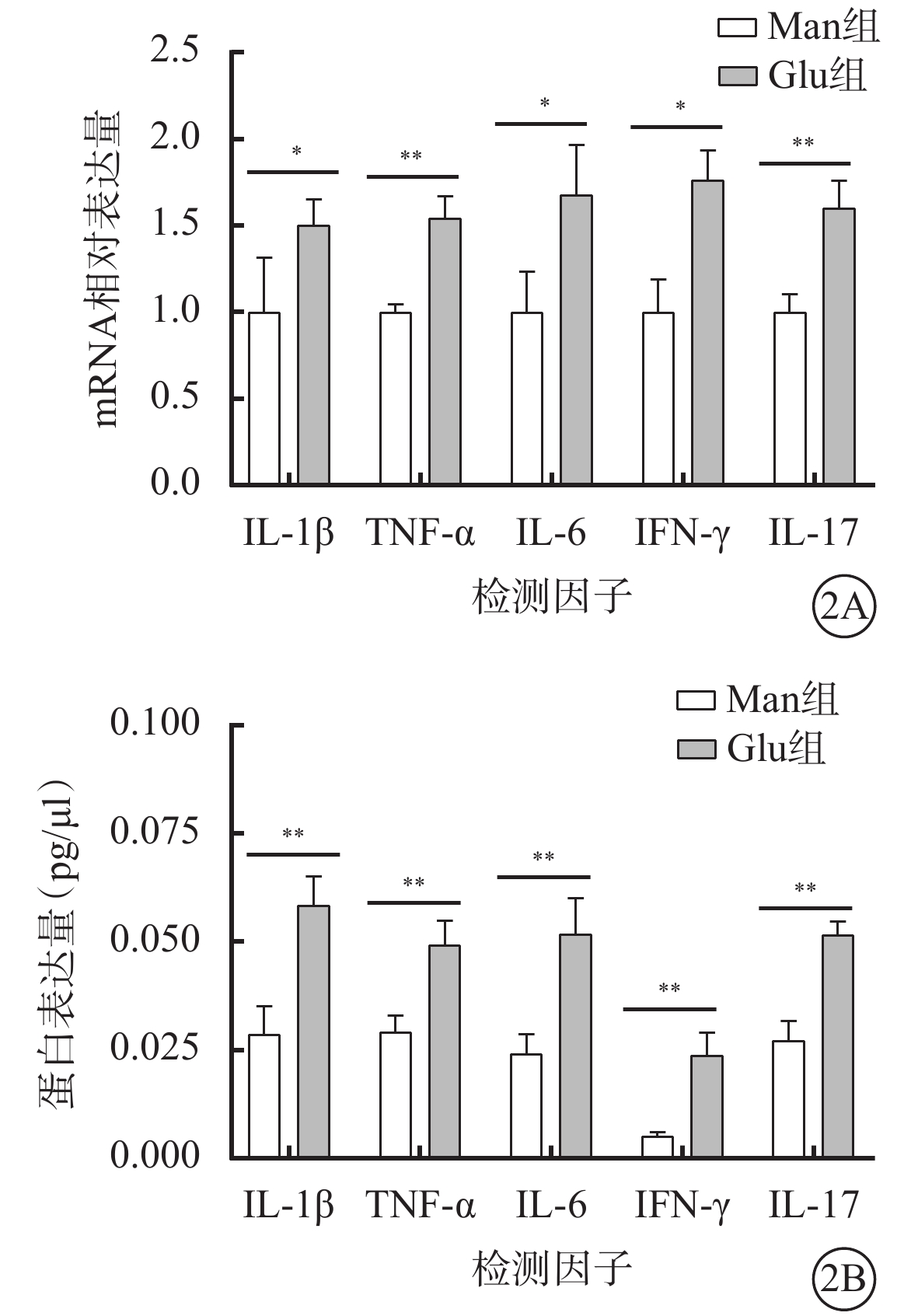
高糖誘導小膠質細胞模型建立成功。免疫熒光染色結果顯示,Man組CD11b強表達,Iba-1不表達(圖1A);Glu組CD11b和Iba-1同時強表達(圖1B)。RT-PCR、AimPlex流式高通量多因子檢測結果顯示,與Man組比較,Glu組細胞中IL-1β、TNF-α、IL-6、IFN-γ、IL-17 mRNA(t=-2.877、-7.057、-3.484、-4.603、-6.396)、蛋白表達量(t=-5.598、-5.177、-5.122、-6.178、-7.727)顯著增加,差異有統計學意義(P<0.05)(圖2)。
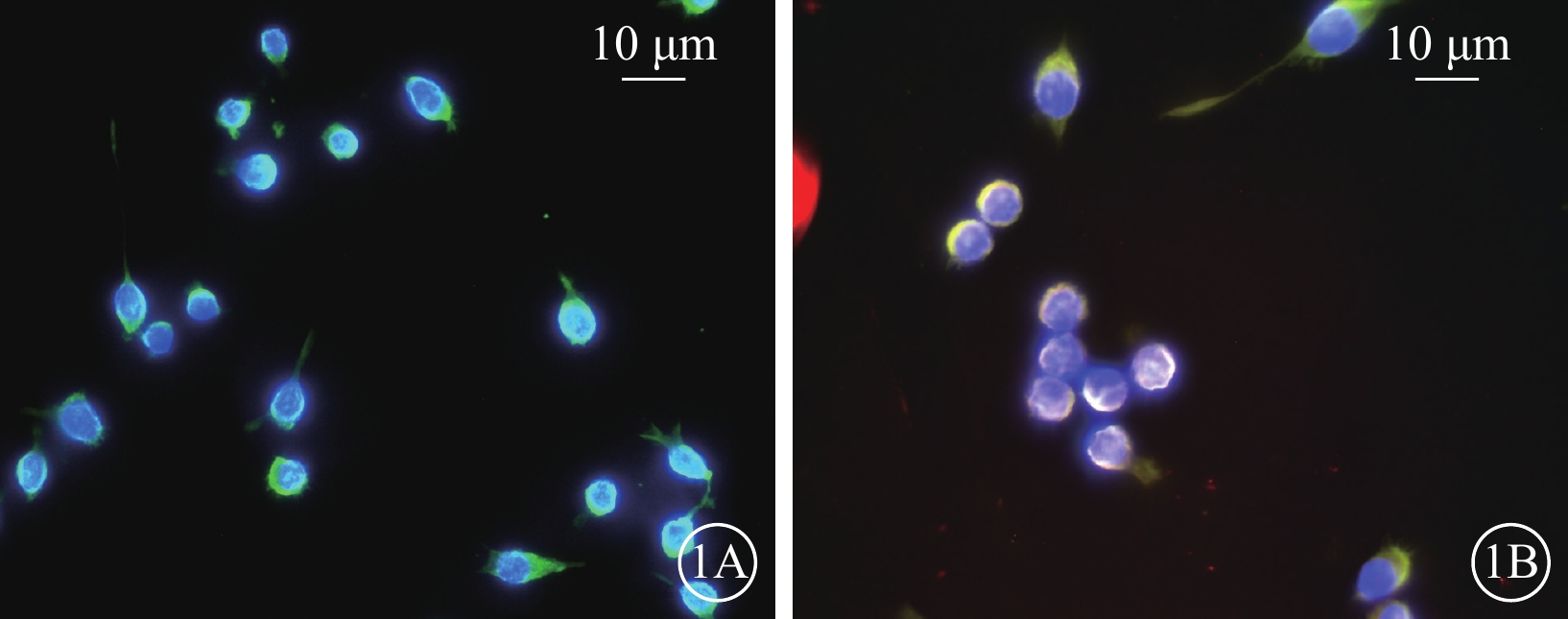 圖1
甘露醇(Man)組、葡萄糖(Glu)組BV2細胞熒光顯微鏡像(n=3) 1A、1B分別示Man組、Glu組。Iba-1標記的BV2細胞顯示為紅色熒光; CD11b標記的BV2細胞顯示為綠色熒光;4',6-二脒基-2-苯基吲哚標記的細胞核顯示為藍色熒光 標尺:10 μm
圖1
甘露醇(Man)組、葡萄糖(Glu)組BV2細胞熒光顯微鏡像(n=3) 1A、1B分別示Man組、Glu組。Iba-1標記的BV2細胞顯示為紅色熒光; CD11b標記的BV2細胞顯示為綠色熒光;4',6-二脒基-2-苯基吲哚標記的細胞核顯示為藍色熒光 標尺:10 μm
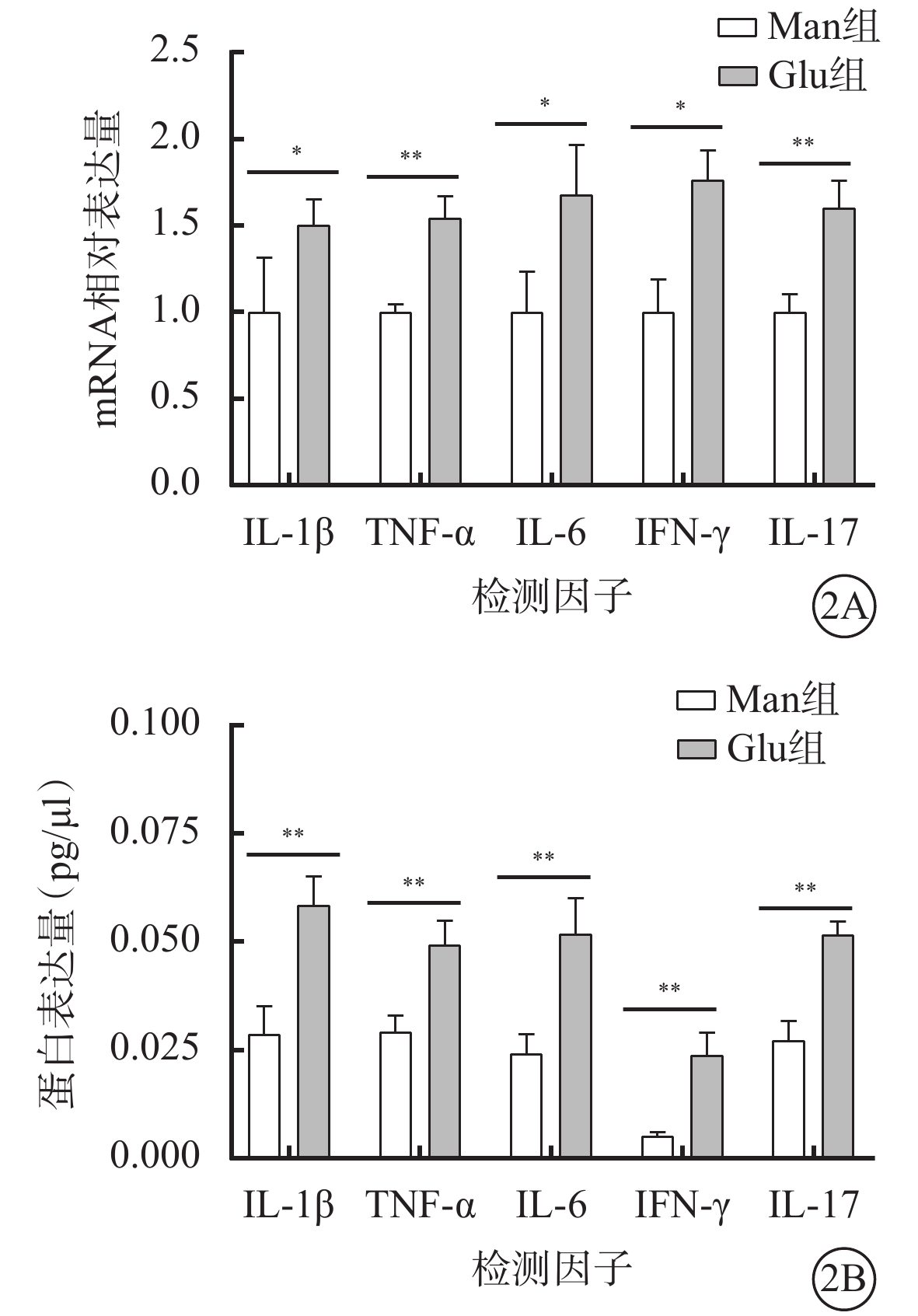 圖2
甘露醇(Man)組、葡萄糖(Glu)組BV2細胞中白細胞介素(IL)-1β、腫瘤壞死因子(TNF)-α、IL-6、干擾素(IFN)-γ、IL-17 mRNA、蛋白表達量比較(n=4) 2A示mRNA相對表達量;2B示蛋白相對表達量。*P<0.05,**P<0.01,***P<0.001
圖2
甘露醇(Man)組、葡萄糖(Glu)組BV2細胞中白細胞介素(IL)-1β、腫瘤壞死因子(TNF)-α、IL-6、干擾素(IFN)-γ、IL-17 mRNA、蛋白表達量比較(n=4) 2A示mRNA相對表達量;2B示蛋白相對表達量。*P<0.05,**P<0.01,***P<0.001
2.2 重組慢病毒對BV2細胞中TAGLN2基因的沉默和過表達
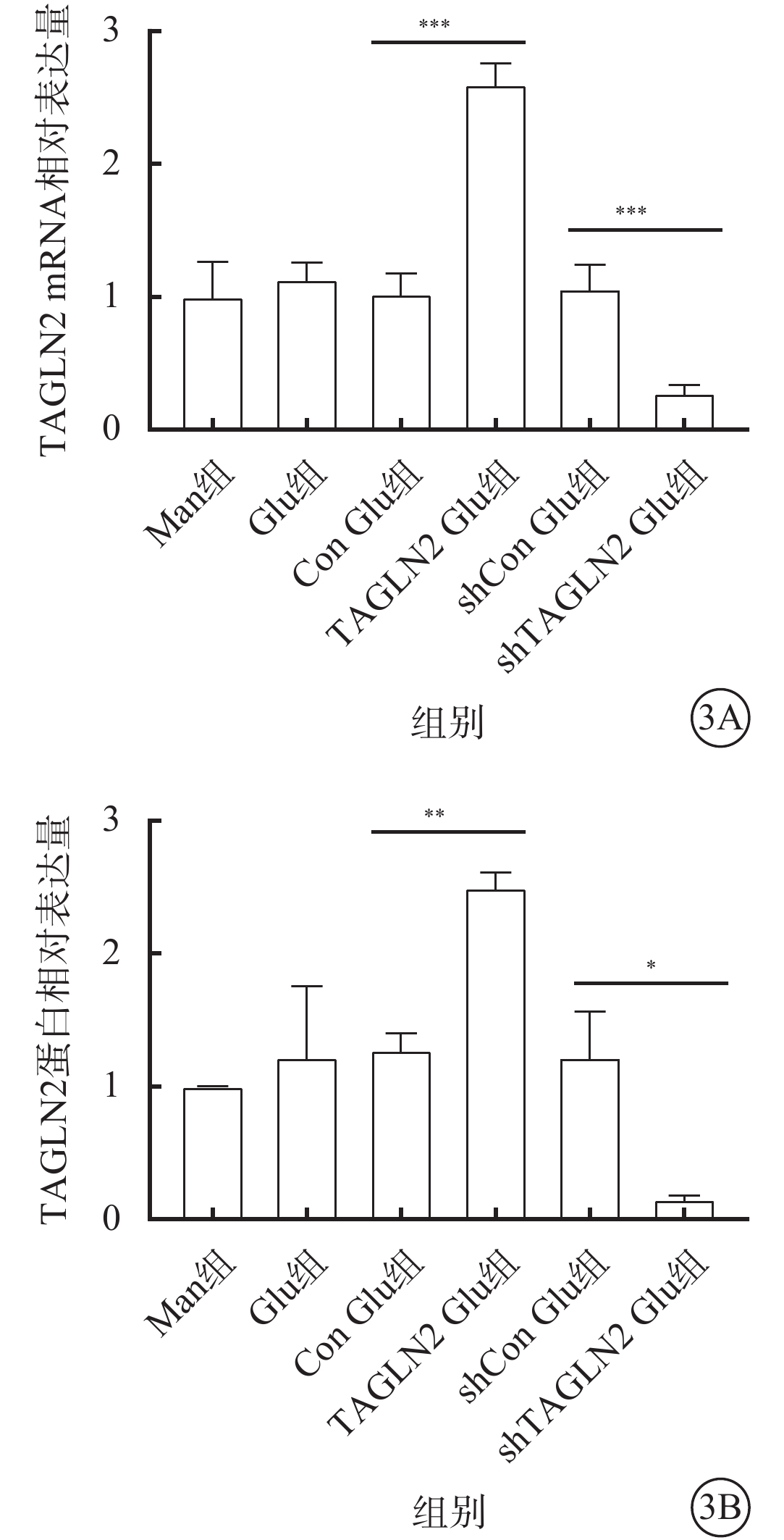
與Con Glu組比較,TAGLN2 Glu組細胞中TAGLN2 mRNA、蛋白相對表達量均顯著增加,差異有統計學意義(t=-14.38、-10.43,P<0.001、0.01),過表達TAGLN2的穩轉BV2細胞系構建成功;與shCon Glu組比較,shTAGLN2 Glu組細胞中TAGLN2 mRNA、蛋白相對表達量均顯著降低,差異有統計學意義(t=7.979、4.394,P<0.001)(圖3),沉默TAGLN2的穩轉BV2細胞系構建成功。
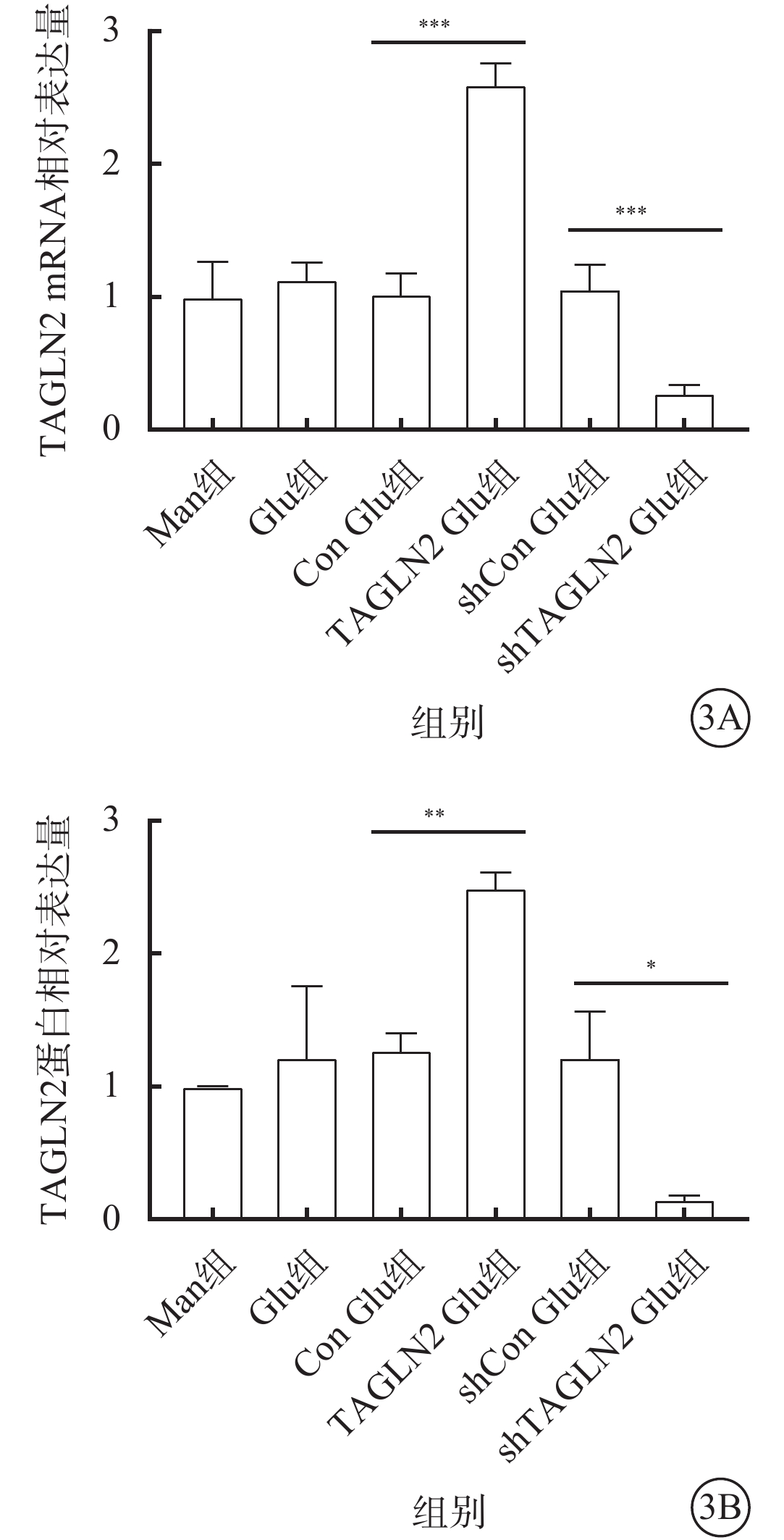 圖3
甘露醇(Man)組、葡萄糖(Glu)組、過表達對照(Con)Glu組、過表達TAGLN2 Glu組、沉默Con Glu(shCon Glu)組、沉默TAGLN2 Glu(shTAGLN2 Glu)組細胞中TAGLN2 mRNA和蛋白相對表達量比較(n=3) 3A示mRNA相對表達量,***P<0.001;3B示蛋白相對表達量,*P<0.05,**P<0.01
圖3
甘露醇(Man)組、葡萄糖(Glu)組、過表達對照(Con)Glu組、過表達TAGLN2 Glu組、沉默Con Glu(shCon Glu)組、沉默TAGLN2 Glu(shTAGLN2 Glu)組細胞中TAGLN2 mRNA和蛋白相對表達量比較(n=3) 3A示mRNA相對表達量,***P<0.001;3B示蛋白相對表達量,*P<0.05,**P<0.01
2.3 DEG表達分析
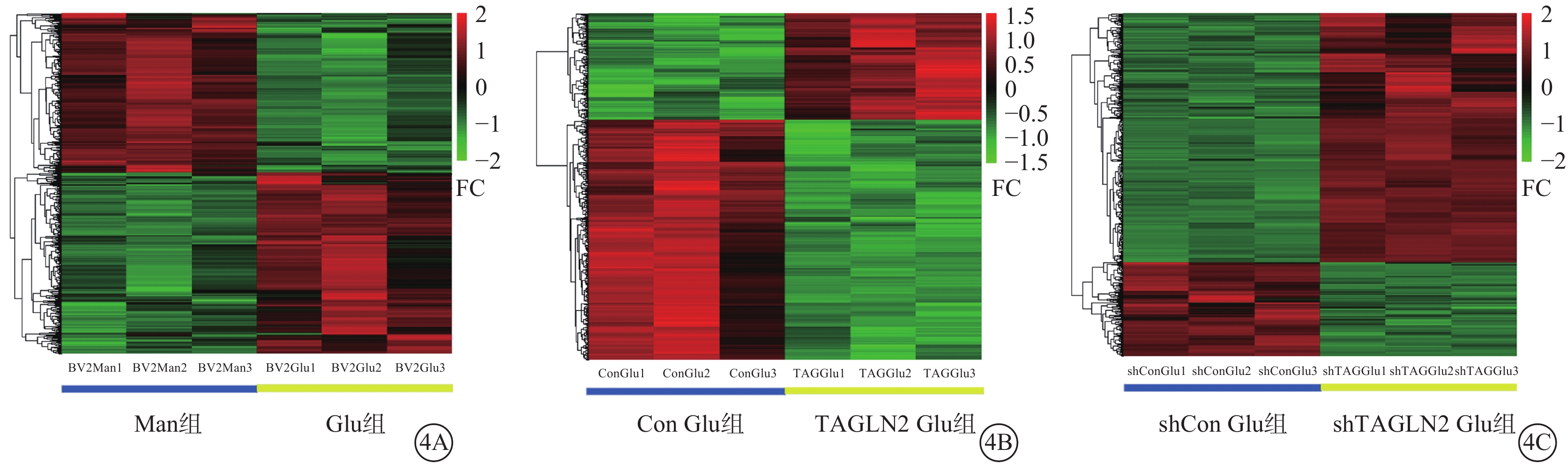
與Man組比較,Glu組篩選出517個DEG,其中上調、下調基因分別為277、240個(圖4A);與Con Glu組比較,TAGLN2 Glu組共篩選480個DEG,其中上調基因、下調基因分別為147、333個(圖4B); 與shCon組比較,shTAGLN2組共篩選 582個DEG,其中上調、下調基因分別為423、159個(圖4C)。
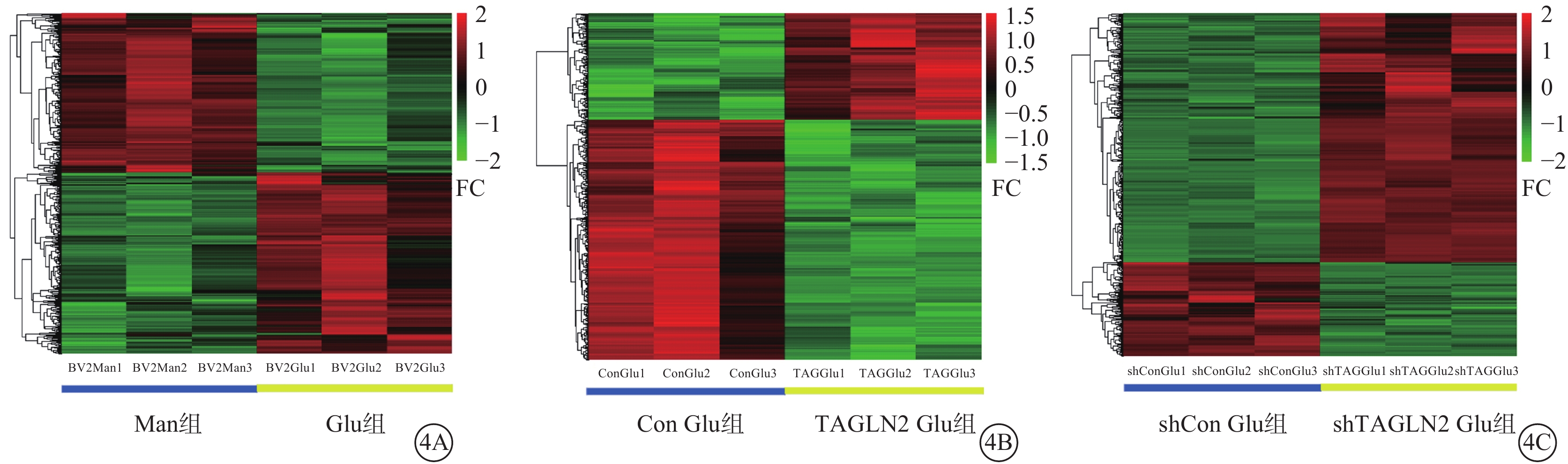 圖4
差異表達基因分析聚類熱圖 4A示甘露醇(Man)組與葡萄糖(Glu)組;4B示過表達對照(Con)Glu組與過表達轉膠蛋白-2(TAGLN2)Glu組;4C示沉默Con Glu(shCon Glu)組與沉默TAGLN2 Glu(shTAGLN2 Glu)組。紅色越深表示差異表達倍數(FC)越高,綠色越深表示FC越低
圖4
差異表達基因分析聚類熱圖 4A示甘露醇(Man)組與葡萄糖(Glu)組;4B示過表達對照(Con)Glu組與過表達轉膠蛋白-2(TAGLN2)Glu組;4C示沉默Con Glu(shCon Glu)組與沉默TAGLN2 Glu(shTAGLN2 Glu)組。紅色越深表示差異表達倍數(FC)越高,綠色越深表示FC越低
2.4 DEG的GO功能富集分析
與Man組相比,Glu組篩選出277個上調DEG顯著富集380條GO條目(P<0.05),主要富集在非經典NF-κB誘導激酶K/NF-κB信號通路及IL-8產生的調控等免疫進程,對氧化應激、炎癥、未折疊蛋白、細胞氧化劑、輻射等刺激應答進程;240個下調DEG顯著富集533條GO條目(P<0.05),主要富集在賴氨酸和蛋白質乙酰化、對轉錄的調控、細胞周期G1/S期轉變的調控、腎小管上皮細胞分化、脂肪細胞分化、橫紋肌細胞分化、神經系統和破骨細胞發育等生物進程。
與Con Glu組相比,TAGLN2 Glu組的147個上調DEG顯著富集392條GO條目(P<0.05),主要富集在脂類、Glu、脂肪酸、甘油醛-3-磷酸生物、三酰甘油、糖原異生和氧化還原等代謝進程;333個下調DEG顯著富集429條GO條目(P<0.05),主要富集在T細胞和B細胞受體信號通路、主要組織相容性復合體(MHC)Ⅱ類分子的抗原加工和提呈、IFN-γ產生等免疫系統進程。
與shCon Glu組相比,shTAGLN2 Glu組的423個上調DEG顯著富集394條GO條目(P<0.05),主要富集在信號素-叢蛋白信號通路、軸突引導中的軸突延伸負調控、鈣離子輸入和轉運、鞘氨醇代謝和脂質磷酸化等進程;159個下調DEG顯著富集333條GO條目(P<0.05),主要富集在對病毒、細菌、IFN-γ、IFN-α和脂多糖應答等先天性免疫應答進程。
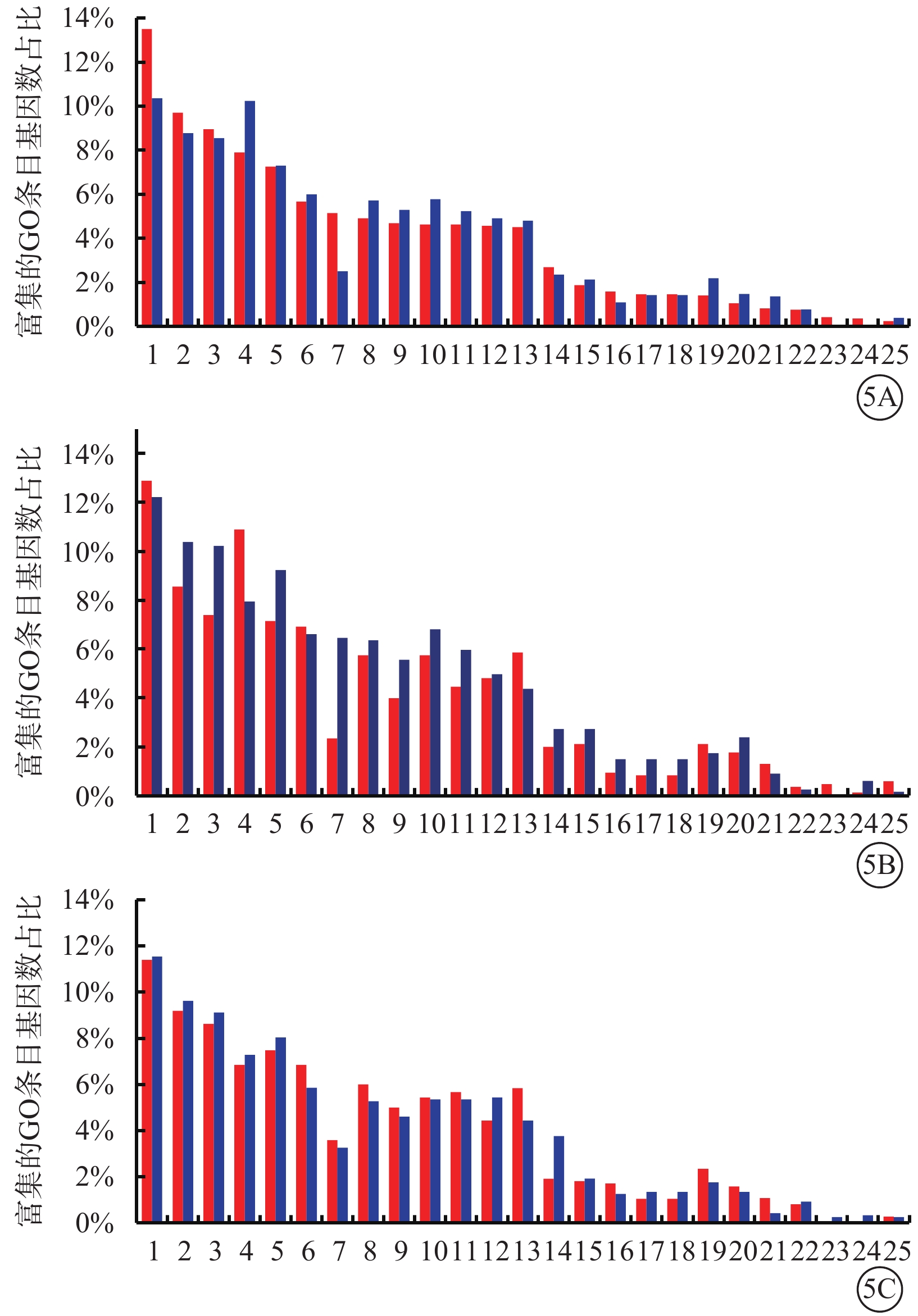
GO生物進程分類分析結果顯示,Glu組與Man組相比,免疫系統進程上調基因較下調基因占比增多,代謝進程上調基因較下調基因占比減少(圖5A);TAGLN2 Glu組與Con Glu組相比,免疫系統進程上調基因比下調基因占比減少,代謝進程上調基因較下調基因占比增多(圖5B);shTAGLN2 Glu組與shCon Glu組相比,上調基因和下調基因在免疫系統進程與代謝進程中占比無顯著變化(圖5C)。
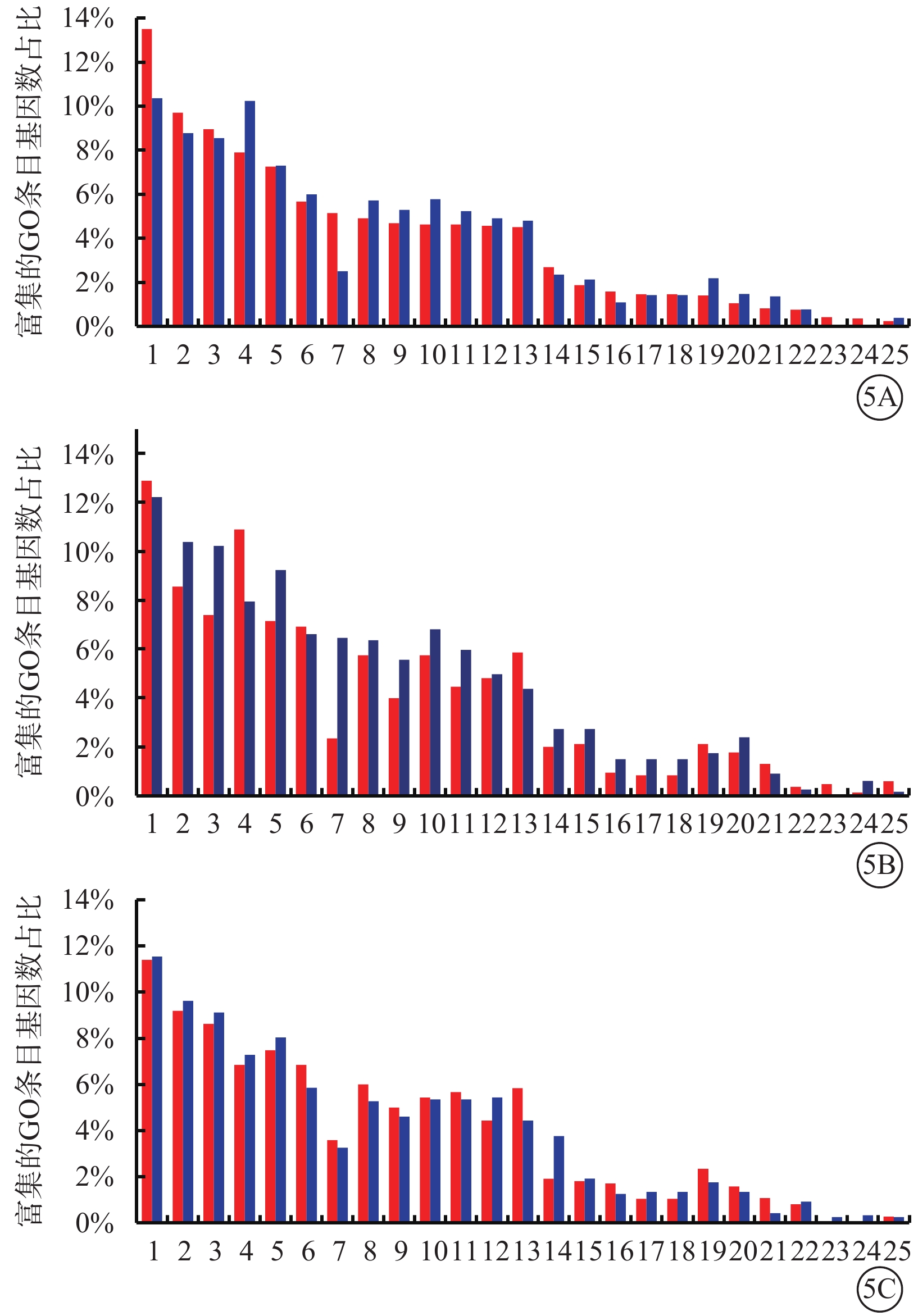 圖5
差異表達基因的基因注釋(GO)生物進程分類分析圖 1:細胞進程;2:生物調節;3:生物進程調控;4:代謝進程;5:對刺激的應答;6:多細胞組織進程;7:免疫系統進程;8:發展過程;9:細胞成分組織或生物發生;10:生物進程正調控;11:信號;12:生物進程負調控;13:定位;14:多組織進程 ;15:細胞增生;16:生物進程;17:生殖;18:生殖進程;19:移動;20:生物粘附;21:生長;22:行為;23:解毒作用;24:細胞殺傷;25:節律進程。5A示葡萄糖(Glu)組與甘露醇(Man)組;5B示過表達轉膠蛋白-2(TAGLN2)Glu組與過表達對照(Con)Glu組;5C示沉默TAGLN2 Glu(shTAGLN2 Glu)組與沉默Con Glu(shCon Glu)組。紅色條柱代表上調基因富集的GO條目基因數占比,藍色條柱代表下調基因富集的GO條目基因數占比
圖5
差異表達基因的基因注釋(GO)生物進程分類分析圖 1:細胞進程;2:生物調節;3:生物進程調控;4:代謝進程;5:對刺激的應答;6:多細胞組織進程;7:免疫系統進程;8:發展過程;9:細胞成分組織或生物發生;10:生物進程正調控;11:信號;12:生物進程負調控;13:定位;14:多組織進程 ;15:細胞增生;16:生物進程;17:生殖;18:生殖進程;19:移動;20:生物粘附;21:生長;22:行為;23:解毒作用;24:細胞殺傷;25:節律進程。5A示葡萄糖(Glu)組與甘露醇(Man)組;5B示過表達轉膠蛋白-2(TAGLN2)Glu組與過表達對照(Con)Glu組;5C示沉默TAGLN2 Glu(shTAGLN2 Glu)組與沉默Con Glu(shCon Glu)組。紅色條柱代表上調基因富集的GO條目基因數占比,藍色條柱代表下調基因富集的GO條目基因數占比
2.5 DEG的KEGG通路富集分析
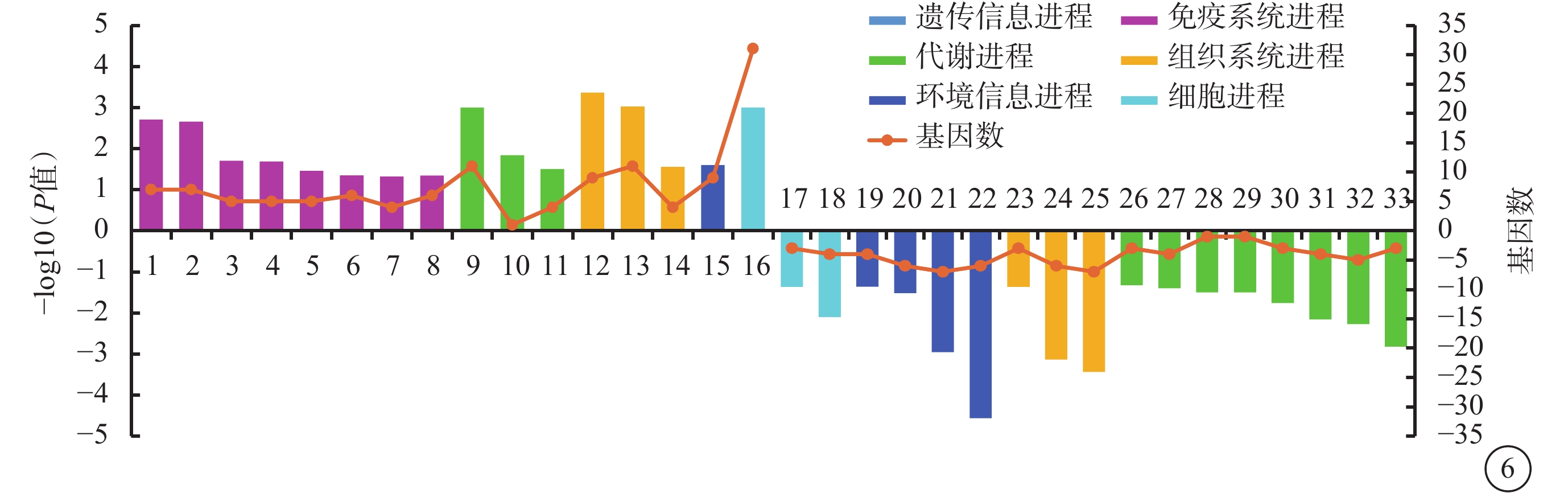
與Man組相比,Glu組上調DEG顯著富集16條KEGG條目(P<0.05),主要與NF-κB、Jak-信號轉導和轉錄激活因子(STAT)、 IL-17信號通路等免疫系統進程相關;下調DEG顯著富集17條KEGG條目(P<0.05),主要與糖胺多聚糖、賴氨酸、膽固醇等代謝進程相關(圖6)。
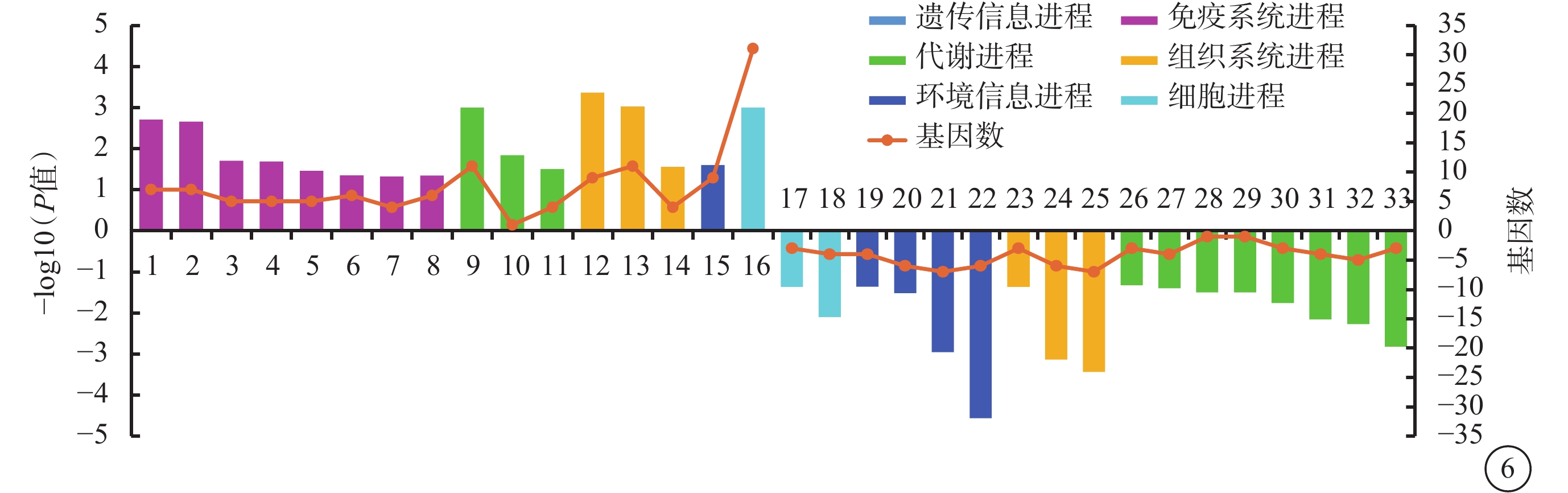 圖6
葡萄糖組與甘露醇組差異表達基因生物進程顯著富集程度前20的京都基因與基因組百科全書條目1:核轉錄因子-κB信號通路;2:蛋白酪氨酸激酶- 信號傳導及轉錄激活蛋白信號通路;3:Toll樣受體信號通路;4:白細胞介素-17信號通路;5:腫瘤壞死因子信號通路;6:趨化因子信號通路;7:輔助性T 細胞17細胞分化;8:核苷酸寡聚化結構域樣受體信號通路;9:氧化磷酸化;10:C5-支二元酸代謝;11:谷胱甘肽代謝;12:逆行內源性大麻素信號;13:生熱作用;14:心肌收縮;15:磷脂酰肌醇三羥基激酶-蛋白激酶B信號通路;16:核糖體;17:線粒體自噬;18:粘附連接;19:低氧誘導因子-1信號通路;20:腎素-血管緊張素系統相關蛋白1信號通路;21:Wnt信號通路;22:Notch信號通路;23:長時程增強;24:黑素原生成;25:甲狀腺激素信號通路;26:甘油脂代謝;27:甲狀旁腺激素的合成和分泌;28:檸檬烯和蒎烯的降解;29:昆蟲激素的生物合成;30:膽固醇代謝;31:賴氨酸降解;32:醛固酮的合成和分泌;33:糖胺多聚糖的降解。橫軸左、右側柱狀圖分別代表上調基因、下調基因,折線圖代表該條目包含的基因數
圖6
葡萄糖組與甘露醇組差異表達基因生物進程顯著富集程度前20的京都基因與基因組百科全書條目1:核轉錄因子-κB信號通路;2:蛋白酪氨酸激酶- 信號傳導及轉錄激活蛋白信號通路;3:Toll樣受體信號通路;4:白細胞介素-17信號通路;5:腫瘤壞死因子信號通路;6:趨化因子信號通路;7:輔助性T 細胞17細胞分化;8:核苷酸寡聚化結構域樣受體信號通路;9:氧化磷酸化;10:C5-支二元酸代謝;11:谷胱甘肽代謝;12:逆行內源性大麻素信號;13:生熱作用;14:心肌收縮;15:磷脂酰肌醇三羥基激酶-蛋白激酶B信號通路;16:核糖體;17:線粒體自噬;18:粘附連接;19:低氧誘導因子-1信號通路;20:腎素-血管緊張素系統相關蛋白1信號通路;21:Wnt信號通路;22:Notch信號通路;23:長時程增強;24:黑素原生成;25:甲狀腺激素信號通路;26:甘油脂代謝;27:甲狀旁腺激素的合成和分泌;28:檸檬烯和蒎烯的降解;29:昆蟲激素的生物合成;30:膽固醇代謝;31:賴氨酸降解;32:醛固酮的合成和分泌;33:糖胺多聚糖的降解。橫軸左、右側柱狀圖分別代表上調基因、下調基因,折線圖代表該條目包含的基因數
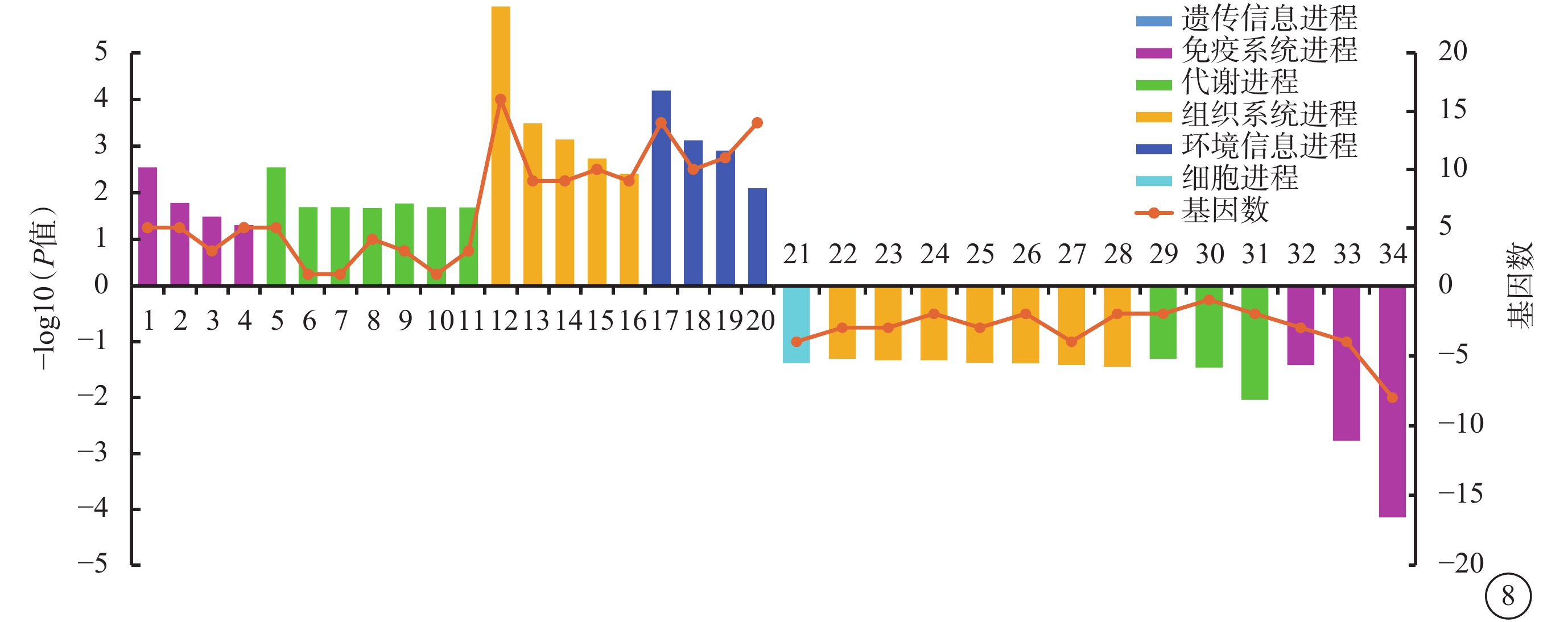
與Con Glu組相比,TAGLN2 Glu組上調DEG顯著富集44條KEGG條目(P<0.05),主要與碳、脂肪酸、甘油脂、丙酮酸、氯代烷和氯烯烴、氨基酸等代謝進程相關;下調DEG顯著富集25條KEGG條目(P<0.05),主要與T細胞與B細胞受體信號通路,Th1、Th2、Th17細胞分化,腸道免疫網絡與IgA產生,NF-κB、TNF、Jak-STAT、趨化因子信號通路以及白細胞、跨內皮細胞遷移等免疫系統進程相關(圖7)。
 圖7
過表達轉膠蛋白-2葡萄糖組與過表達對照葡萄糖組差異表達基因生物進程顯著富集程度前20的京都基因與基因組百科全書條目 1:不同環境中的微生物代謝;2:代謝通路;3:碳代謝;4:脂肪酸降解;5:脂肪酸代謝;6:甘油脂代謝;7:丙酮酸代謝;8:氯代烷和氯烯烴的降解;9:糖酵解/糖異生;10:纈氨酸、亮氨酸和異亮氨酸的降解;11:氨基酸的生物合成;12:次生代謝物的生物合成;13:抗生素的生物合成;14:生熱作用;15:胰島素信號通路;1:6:脂肪細胞脂解作用的調控;17:近端小管碳酸氫鹽回收;18:過氧化物酶體增生物激活受體信號通路;19:腺苷5'-單磷酸激活蛋白激酶信號通路;20過氧物酶體;21:緊密連接;22:灶性粘連;23:腎素-血管緊張素系統信號通路;24:細胞粘附分子;25:長時程增強;26:血小板活化;27:鞘糖脂的生物合成;28:趨化因子信號通路;29: Fc段γ受體介導的吞噬作用;30:蛋白酪氨酸激酶- 信號傳導及轉錄激活蛋白信號通路信號通路;31:原處理和提呈;32:腫瘤壞死因子信號通路;33:白細胞跨內皮細胞遷移;34:核因子-κB信號通路;35:腸道免疫網絡與免疫球蛋白A的產生;36:輔助性T細胞(Th)17細胞分化;37:B細胞受體信號通路;38:Th1和Th2細胞分化;39:T細胞受體信號通路;40:造血細胞譜系。橫軸左、右側柱狀圖分別代表上調基因、下調基因,折線圖代表該條目包含的基因數
圖7
過表達轉膠蛋白-2葡萄糖組與過表達對照葡萄糖組差異表達基因生物進程顯著富集程度前20的京都基因與基因組百科全書條目 1:不同環境中的微生物代謝;2:代謝通路;3:碳代謝;4:脂肪酸降解;5:脂肪酸代謝;6:甘油脂代謝;7:丙酮酸代謝;8:氯代烷和氯烯烴的降解;9:糖酵解/糖異生;10:纈氨酸、亮氨酸和異亮氨酸的降解;11:氨基酸的生物合成;12:次生代謝物的生物合成;13:抗生素的生物合成;14:生熱作用;15:胰島素信號通路;1:6:脂肪細胞脂解作用的調控;17:近端小管碳酸氫鹽回收;18:過氧化物酶體增生物激活受體信號通路;19:腺苷5'-單磷酸激活蛋白激酶信號通路;20過氧物酶體;21:緊密連接;22:灶性粘連;23:腎素-血管緊張素系統信號通路;24:細胞粘附分子;25:長時程增強;26:血小板活化;27:鞘糖脂的生物合成;28:趨化因子信號通路;29: Fc段γ受體介導的吞噬作用;30:蛋白酪氨酸激酶- 信號傳導及轉錄激活蛋白信號通路信號通路;31:原處理和提呈;32:腫瘤壞死因子信號通路;33:白細胞跨內皮細胞遷移;34:核因子-κB信號通路;35:腸道免疫網絡與免疫球蛋白A的產生;36:輔助性T細胞(Th)17細胞分化;37:B細胞受體信號通路;38:Th1和Th2細胞分化;39:T細胞受體信號通路;40:造血細胞譜系。橫軸左、右側柱狀圖分別代表上調基因、下調基因,折線圖代表該條目包含的基因數
與shCon Glu組相比,shTAGLN2 Glu組上調DEG顯著富集40條KEGG條目(P<0.05),主要與TNF、趨化因子信號通路等免疫進程及色氨酸、環氧氯己烷、鞘脂、鞘糖脂和丁酸等代謝進程相關;下調DEG顯著富集14條KEGG條目(P<0.05),主要與核苷酸寡聚化結構域樣受體、Toll樣受體信號通路以及氮代謝、脂肪消化吸收等進程相關(圖8)。
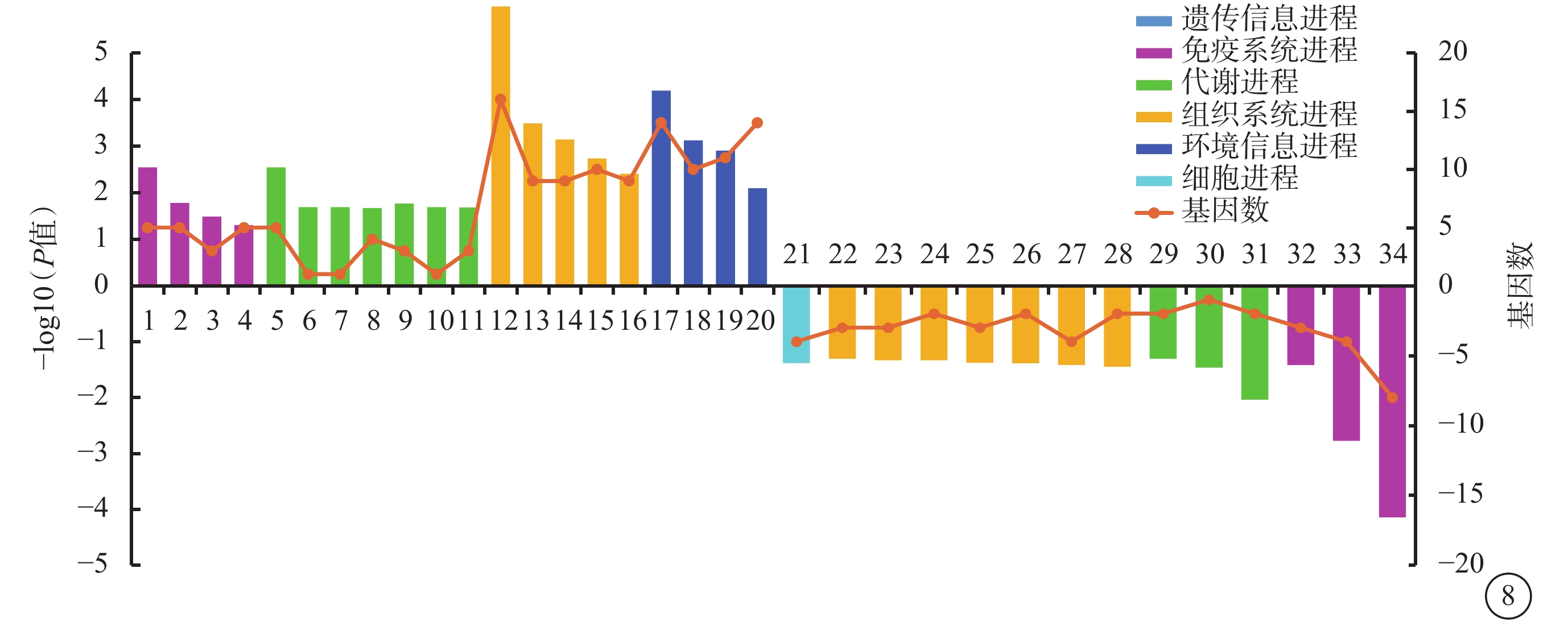 圖8
沉默轉膠蛋白-2葡萄糖組與沉默對照葡萄糖組差異表達基因生物進程顯著富集程度前20的京都基因與基因組百科全書條目 1:腫瘤壞死因子信號通路;2:趨化因子信號通路;3:輔助性T細胞(Th)1和Th2細胞分化;4:細胞因子-細胞因子受體相互作用;5:色氨酸代謝;6:環氧氯己烷與氯苯的降解;7:氟苯甲酸鹽的降解;8:鞘脂代謝;9:糖脂的生物合成;10:甲苯降解;11:丁酸代謝;12:軸突引導;13:醛固酮的合成和分泌;14:膽堿能突觸;15:催產素信號通路;16:多巴胺能突觸;17:鈣信號通路;18:心血管活性多肽信號通路;19:環磷酸鳥苷-蛋白激酶G信號通路;20:絲裂原活化蛋白激酶信號通路;21:肌動蛋白細胞骨架的調控;22:膽堿能突觸;23:松弛素信號通路;24:胞質DNA傳感途徑;25:破骨細胞分化;26:內分泌和其他因素調節的鈣重吸收;27:軸突引導;28:脂肪消化吸收;29:細胞色素P450對外源物質的代謝作用;30:纈氨酸、亮氨酸和異亮氨酸的生物合成;31:氮代謝;32:Toll樣受體信號通路;33:視黃酸(維甲酸)誘導基因蛋白-I樣受體信號通路;34:核苷酸寡聚化結構域樣受體信號通路。橫軸左、右側柱狀圖分別代表上調基因、下調基因,折線圖代表該條目包含的基因數
圖8
沉默轉膠蛋白-2葡萄糖組與沉默對照葡萄糖組差異表達基因生物進程顯著富集程度前20的京都基因與基因組百科全書條目 1:腫瘤壞死因子信號通路;2:趨化因子信號通路;3:輔助性T細胞(Th)1和Th2細胞分化;4:細胞因子-細胞因子受體相互作用;5:色氨酸代謝;6:環氧氯己烷與氯苯的降解;7:氟苯甲酸鹽的降解;8:鞘脂代謝;9:糖脂的生物合成;10:甲苯降解;11:丁酸代謝;12:軸突引導;13:醛固酮的合成和分泌;14:膽堿能突觸;15:催產素信號通路;16:多巴胺能突觸;17:鈣信號通路;18:心血管活性多肽信號通路;19:環磷酸鳥苷-蛋白激酶G信號通路;20:絲裂原活化蛋白激酶信號通路;21:肌動蛋白細胞骨架的調控;22:膽堿能突觸;23:松弛素信號通路;24:胞質DNA傳感途徑;25:破骨細胞分化;26:內分泌和其他因素調節的鈣重吸收;27:軸突引導;28:脂肪消化吸收;29:細胞色素P450對外源物質的代謝作用;30:纈氨酸、亮氨酸和異亮氨酸的生物合成;31:氮代謝;32:Toll樣受體信號通路;33:視黃酸(維甲酸)誘導基因蛋白-I樣受體信號通路;34:核苷酸寡聚化結構域樣受體信號通路。橫軸左、右側柱狀圖分別代表上調基因、下調基因,折線圖代表該條目包含的基因數
2.6 KEGG免疫系統進程和免疫相關DEG的PPI分析
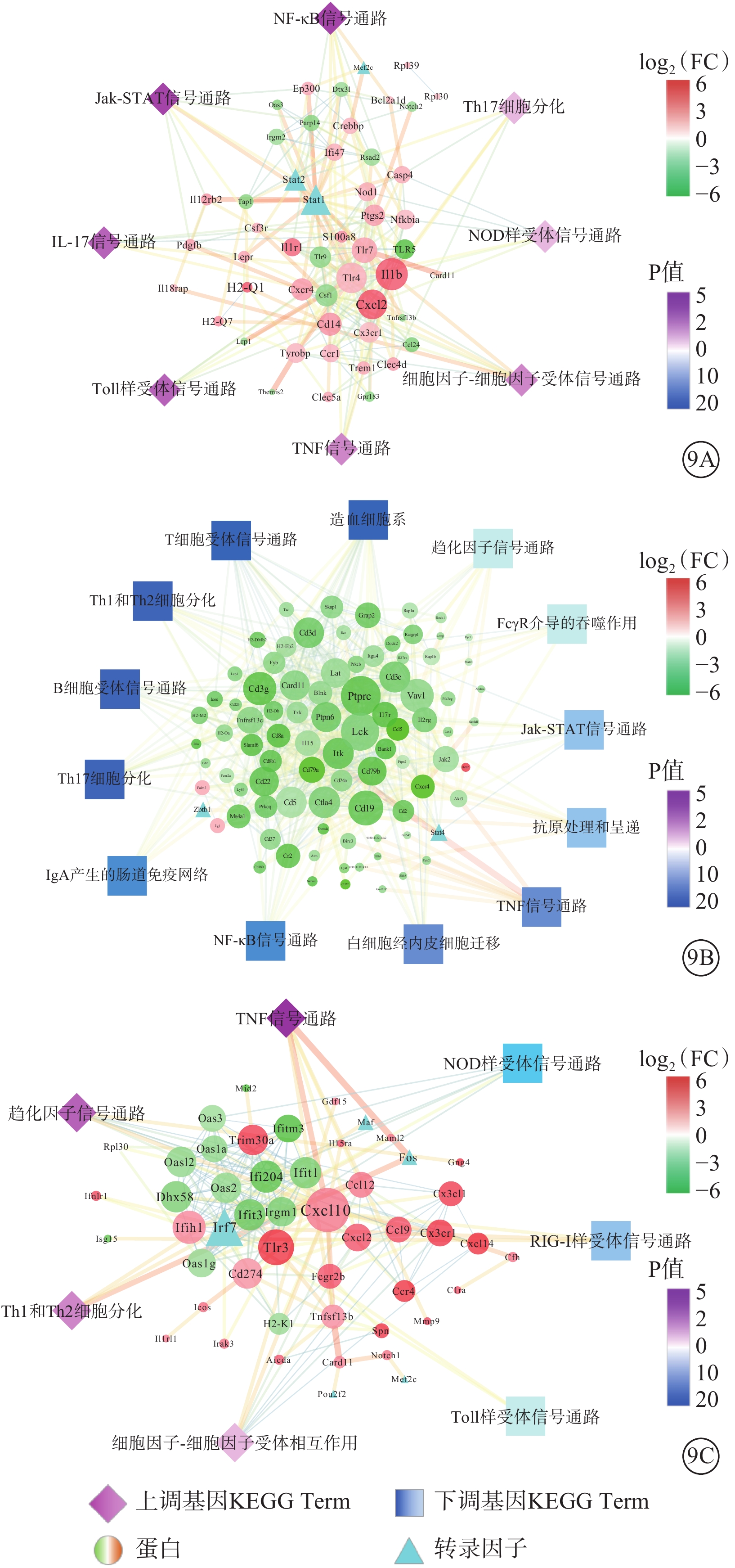
與Man組相比,Glu組有58個與免疫相關DEG,8條免疫系統進程。包括Stat1、Stat2和Mef2c 3個轉錄因子,權重最高的10個DEG分別為Il1b、Stat1、Tlr4、Cxcl2、Tlr7、Cd14、Stat2、Csf1、Cxcr4、Tlr9(圖9A)。
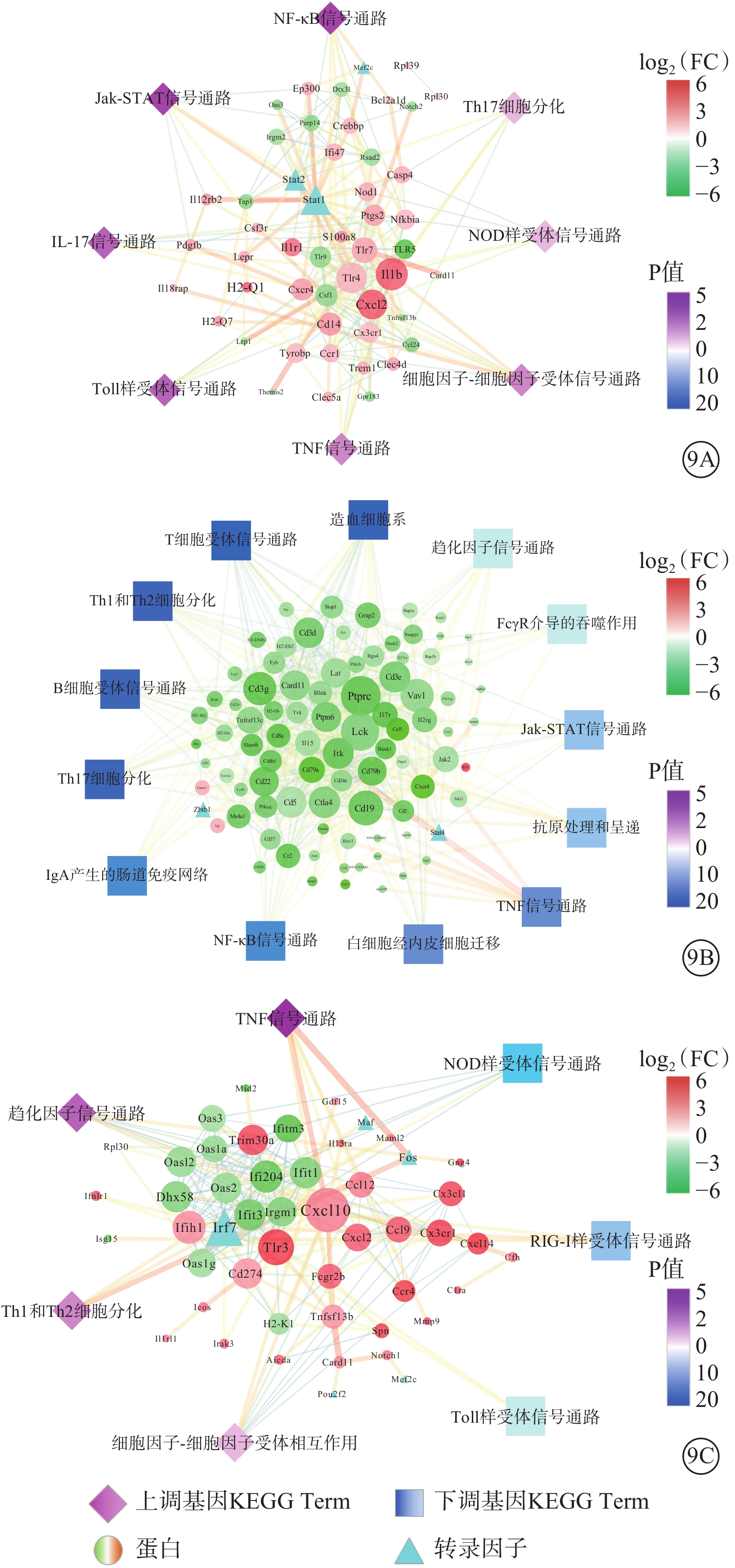 圖9
蛋白-蛋白互作網絡圖 NF:核因子;Jak-STAT:蛋白酪氨酸激酶- 信號傳導及轉錄激活蛋白;IL-白細胞介素;Th:輔助性T細胞;NOD:核苷酸寡聚化結構域;FcγR:Fc段γ受體;IgA:免疫球蛋白A;TNF:腫瘤壞死因子;RIG:視黃酸(維甲酸)誘導基因蛋白;9A示葡萄糖(Glu)組與甘露醇(Man)組;9B示過表達轉膠蛋白-2(TAGLN2)Glu組與過表達對照(Con)Glu組;7C示沉默TAGLN2 Glu(shTAGLN2 Glu)組與沉默Con Glu(shCon Glu)組。紅色越深表示差異基因表達量越高,綠色越深表示差異基因表達量越低;藍色、紫色越深表示P值越大
圖9
蛋白-蛋白互作網絡圖 NF:核因子;Jak-STAT:蛋白酪氨酸激酶- 信號傳導及轉錄激活蛋白;IL-白細胞介素;Th:輔助性T細胞;NOD:核苷酸寡聚化結構域;FcγR:Fc段γ受體;IgA:免疫球蛋白A;TNF:腫瘤壞死因子;RIG:視黃酸(維甲酸)誘導基因蛋白;9A示葡萄糖(Glu)組與甘露醇(Man)組;9B示過表達轉膠蛋白-2(TAGLN2)Glu組與過表達對照(Con)Glu組;7C示沉默TAGLN2 Glu(shTAGLN2 Glu)組與沉默Con Glu(shCon Glu)組。紅色越深表示差異基因表達量越高,綠色越深表示差異基因表達量越低;藍色、紫色越深表示P值越大
與Con Glu組相比,TAGLN2 Glu組有104個與免疫相關DEG,13條免疫系統進程。包括Stat4、Zbtb1 2個轉錄因子,權重最高的10個DEG分別是Ptprc、Lck、Cd19、Vav1、Cd3g、Ctla4、Cd5、Cd3e、Itk、Cd3d(圖9B)。
與shCon Glu組相比,shTAGLN2 Glu組有68個與免疫相關DEG,7條免疫系統進程,包括Fos、Pou2f2、Maf、Mef2c和Irf7等5個轉錄因子,權重最高的10個DEG分別是Cxcl10、Irf7、Tlr3、Ifih1、Ifi204、Ifit1、Ifit3、Dhx58、Cd274、Trim30a(圖9C)。
2.7 TAGLN2抑制高糖誘導的炎癥因子表達
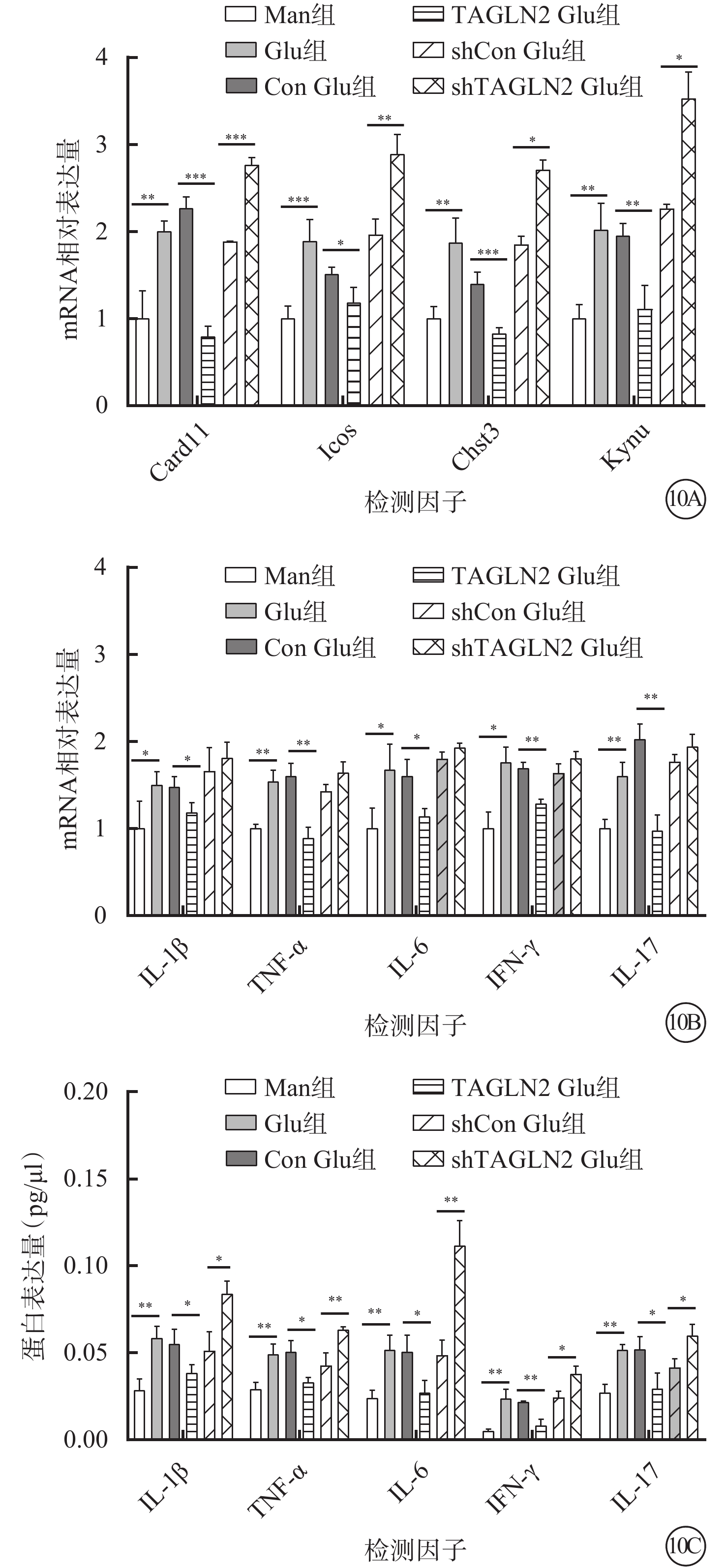
RT-PCR檢測結果顯示,與Man組比較,Glu組細胞中Card11、Icos、Chst3、Kynu mRNA相對表達量顯著增高,差異有統計學意義(t=-5.925、-6.339、-5.523、-5.948,P=0.001、0.001、0.003、0.001)。與Con Glu組比較,TAGLN2 Glu組中Card11、Icos、Chst3、Kynu mRNA相對表達量顯著降低,差異有統計學意義(t=13.530、3.482、6.949、5.399,P=0.000、0.018、0.001、0.006)(圖10A)。
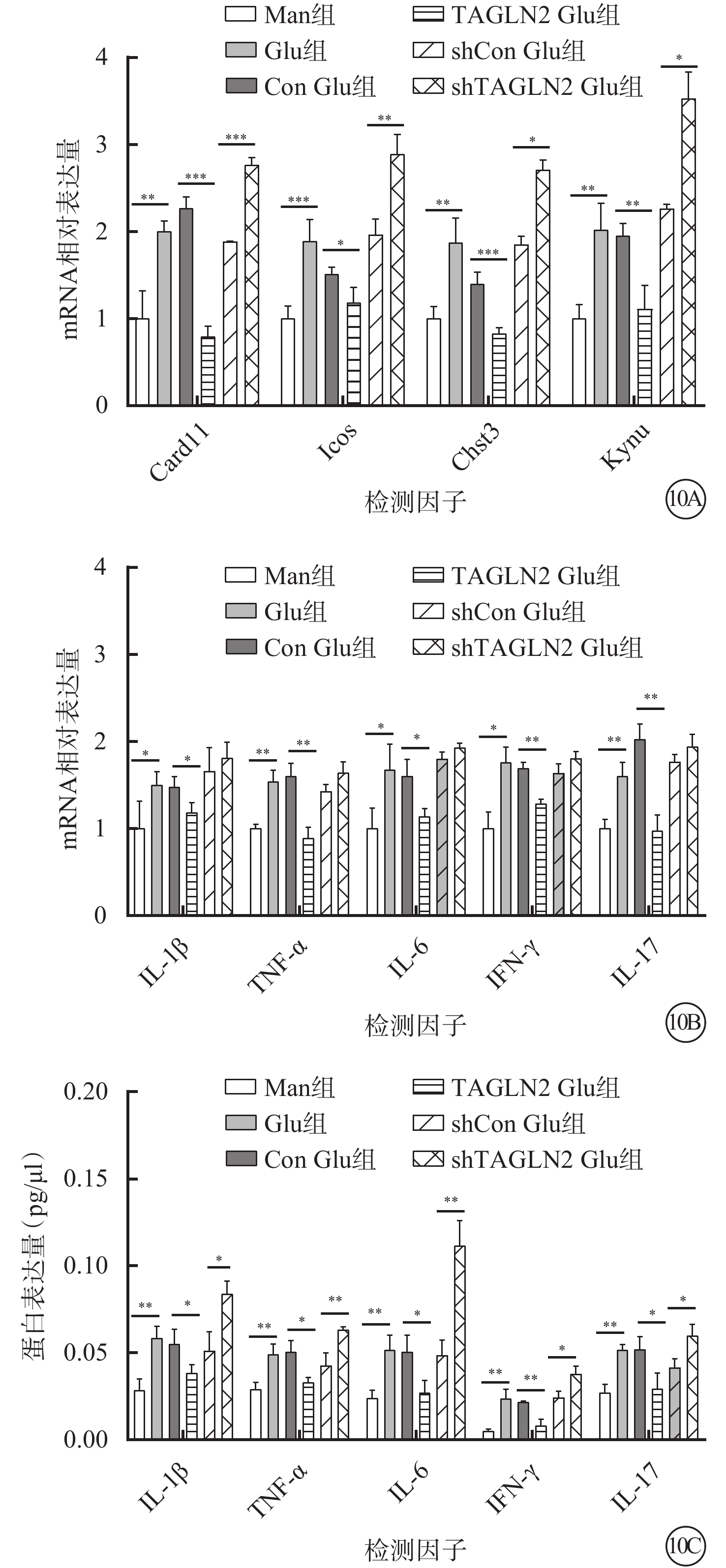 圖10
甘露醇(Man)組、葡萄糖(Glu)組、過表達對照(Con)Glu組、過表達TAGLN2 Glu組、沉默Con Glu(shCon Glu)組、沉默TAGLN2 Glu(shTAGLN2 Glu)組細胞中各因子mRNA和蛋白相對表達量比較(n=3) 10A示Card11、Icos、Chst3、Kynu mRNA相對表達量,*P<0.05,**P<0.01,***P<0.001;10B示白細胞介素(IL)-1β、腫瘤壞死因子(TNF)-α、IL-6、干擾素(IFN)-γ、IL-17 mRNA相對表達量,*P<0.05,**P<0.01;10C示IL-1β、TNF-α、IL-6、IFN-γ、IL-17 蛋白相對表達量,*P<0.05,**P<0.01
圖10
甘露醇(Man)組、葡萄糖(Glu)組、過表達對照(Con)Glu組、過表達TAGLN2 Glu組、沉默Con Glu(shCon Glu)組、沉默TAGLN2 Glu(shTAGLN2 Glu)組細胞中各因子mRNA和蛋白相對表達量比較(n=3) 10A示Card11、Icos、Chst3、Kynu mRNA相對表達量,*P<0.05,**P<0.01,***P<0.001;10B示白細胞介素(IL)-1β、腫瘤壞死因子(TNF)-α、IL-6、干擾素(IFN)-γ、IL-17 mRNA相對表達量,*P<0.05,**P<0.01;10C示IL-1β、TNF-α、IL-6、IFN-γ、IL-17 蛋白相對表達量,*P<0.05,**P<0.01
RT-PCR、AimPlex流式高通量多因子檢測結果顯示,與Con Glu組比較,TAGLN2 Glu組細胞中IL-1β、TNF-α、IL-6、IFN-γ、IL-17 mRNA(t=2.960、5.800、3.130、7.690、6.530)、蛋白相對表達量(t=3.050、4.530、3.420、6.880、3.380)均顯著下降,差異有統計學意義(P<0.05)(圖10B,10C)。
3 討論
既往研究表明,高糖能誘導小膠質細胞NF-κB信號通路激活[3-4],也能誘導視網膜血管內皮細胞和視網膜中Jak-STAT信號通路活化,并釋放大量炎癥因子和趨化因子[12-13]。本研究結果顯示,高糖能夠激活BV2細胞中NF-κB、Jak-STAT信號通路,促進TNF-α、IL-6和IL-1β等多種炎癥因子和趨化因子釋放。研究表明,TAGLN2過表達抑制淋巴毒素-β受體激動劑誘導的血管平滑肌細胞中NF-κB誘導激酶及其下游NF-κB的典型和非典型信號通路的表達[14]。本研究也發現過表達TAGLN2能夠顯著抑制高糖誘導的NF-κB信號通路的激活和炎癥因子、趨化因子的釋放。GO富集分析也發現過表達TAGLN2后小膠質細胞中T細胞和B細胞受體信號通路、MHC Ⅱ類分子的抗原加工和提呈、白細胞與細胞粘附及調節和IFN-γ產生的正向調節等免疫系統進程被顯著抑制。沙門菌感染后引起TAGLN2?/?小鼠的血漿TNF-α、IL-6、IL-1β、IL-12水平顯著升高[9]。本研究結果顯示,沉默TAGLN2后高糖誘導的小膠質細胞中TNF信號通路、趨化因子和細胞因子-細胞因子受體相互作用信號通路被激活,TNF-α、IL-6、IL-17、IL-1β等炎癥因子和趨化因子表達增加,但是Toll樣受體信號通路和視黃酸(維甲酸)誘導基因蛋白-Ⅰ樣受體信號通路等模式識別受體信號通路被抑制;GO富集分析也發現沉默TAGLN2后小膠質細胞對病毒、細菌、IFN-α、IFN-β和脂多糖的固有免疫應答進程減少,表明沉默TAGLN2后小膠質細胞識別抗原和外部刺激能力減弱,炎癥反應增加,免疫功能失衡。因此,TAGLN2在高糖誘導的小膠質細胞炎癥反應中發揮了關鍵作用,是治療DR的潛在藥物靶點。
本研究結果顯示,分別與Man組、Con Glu組、shCon Glu組這3個對照組比較,Glu組、TAGLN2 Glu組及shTAGLN2 Glu組同時表達的DEG為14個,其中Card11、Icos、Chst3、Kynu均與疾病炎癥反應相關。Card11基因參與T細胞受體、B細胞受體介導的NF-κB信號通路活化[15-16]。Icos基因介導非肥胖糖尿病(NOD)小鼠胰島素依賴型糖尿病發展,是NOD小鼠發生胰島素炎和高血糖所不可缺少的;其缺失可導致Th1細胞因子IFN-γ的產生減少[17-18]。抑制Icos/Icos配體信號顯著抑制炎癥因子的表達,以及巨噬細胞、中性和嗜酸性粒細胞浸潤[19-20]。Chst3基因參與硫酸軟骨素生物合成和糖胺聚糖代謝,其沉默可減少肺氣腫巨噬細胞的積累并抑制基質金屬蛋白酶-9的表達,促進疾病恢復[21]。Kynu是牛皮癬和其他炎性疾病中的新型炎性因子[22],抑制小膠質細胞中的Kynu通路能夠保護反應性小膠質細胞相關神經元的萎縮[23]。本研究結果顯示,過表達TAGLN2能夠顯著抑制高糖誘導的小膠質細胞中Card11、Icos、Chst3、Kynu的表達,減少NF-κB信號通路活化和多種炎癥因子和趨化因子的表達。
本研究存在的不足:(1)DR發病機制復雜,高血糖僅是疾病發生的主要原因之一,高糖干預的細胞模型不能完全模擬體內情況,還需要進一步在體實驗加以驗證;(2)本研究僅檢測了高糖處理24 h顯著DEG及信號通路的改變,其中關鍵信號通路的功能分析驗證及更長時間高糖處理DEG和信號通路的改變,仍需要進一步實驗研究。
視網膜小膠質細胞是視網膜內免疫監測細胞,靜止狀態下其通過吞噬和控制低度炎癥幫助維持視網膜的組織穩態。但高血糖引起的長期組織應激可導致小膠質細胞過度反應,并產生促炎細胞因子和趨化因子引起慢性炎癥[1]。在糖尿病視網膜病變(DR)中小膠質細胞活化導致炎癥增加和免疫代謝紊亂[2]。鏈脲佐菌素誘導的DR模型小鼠,小膠質細胞活化并聚集在視網膜色素上皮層,產生的白細胞介素(IL)-6進一步促進小膠質細胞的募集和腫瘤壞死因子(TNF)-α的釋放,加重血視網膜屏障破壞[3]。通過抑制小膠質細胞引發的視網膜炎癥,減少高糖誘導的細胞外調節蛋白激酶1/2-核因子(NF)-κB通路的激活,能夠減輕DR[4]。轉膠蛋白-2(TAGLN2)為肌動蛋白(actin)細胞骨架結合蛋白,屬于轉膠蛋白超家族[5];其主要定位于細胞胞漿,可通過直接調控相關基因表達而影響細胞表型[6]。TAGLN2在平滑肌細胞及成纖維細胞中高表達,參與血管生成、骨架重塑以及細胞遷移、凋亡及增生等功能調控,能夠調節T細胞激活,在B細胞中也參與了T細胞-B細胞結合物的穩定[5-8]。TAGLN2在脂多糖刺激的巨噬細胞中表達高度上調,并促進巨噬細胞的吞噬[9]。重組TAGLN2蛋白能夠增強樹突狀細胞抑制腫瘤生長和轉移的功能[10]。因此TAGLN2在淋巴細胞、巨噬細胞和樹突狀細胞的免疫反應中都發揮了重要的作用。但是TAGLN2是否在高糖誘導的小膠質細胞免疫反應中起作用及其作用機制鮮見報道。轉錄組測序分析技術為細胞功能研究提供了新的方法[11]。本研究利用慢病毒沉默和過表達BV2小膠質細胞(以下簡稱為BV2細胞)中的TAGLN2基因,通過轉錄組測序和生物信息學分析闡明TANLN2調控高糖誘導的小膠質細胞炎癥反應的分子機制,為DR的治療提供新的策略。現將結果報道如下。
1 材料和方法
1.1 主要材料
BV2細胞(國家實驗細胞資源共享服務平臺)。D-葡萄糖(Glu)、甘露醇(Man)(德國Merck公司),嘌呤霉素、蛋白裂解液、山羊血清(北京索萊寶科技有限公司),胎牛血清(美國Gibco 公司),脂質體Lipofectamine? 3000、Trizol試劑、4',6-二脒基-2-苯基吲哚(DAPI)(美國Invitrogen公司),蛋白酶抑制劑(美國MCE公司)。PrimeScript RT reagentKit with gDNA eraser試劑盒、SYBR Rremix Ex Taq II Kit試劑盒(日本TaKaRa公司),二喹啉甲酸(BCA)蛋白濃度測定試劑盒(北京索萊寶科技有限公司),Premixed Multiplex Kit Mouse Custom 8-Plex Kit試劑盒(北京曠博生物技術股份有限公司)。質粒載體:pHelper1.0、pHelper2.0(上海吉凱基因科技有限公司),psPAX2、PMD2.G(漢恒生物科技有限公司)。抗體:抗TAGLN2兔多克隆抗體、抗CD11b兔多克隆抗體(美國Abcam公司),抗Iba-1羊多克隆抗體(美國Novus公司),辣根過氧化物酶(HRP)標記羊抗兔免疫球蛋白G(IgG)、 硫氰酸熒光素(FITC)標記的羊抗兔IgG(FITC-IgG)(中國博奧森生物技術有限公司),藻紅蛋白(PE)標記的驢抗羊IgG[PE-IgG,愛必信(上海)生物科技有限公司]。儀器設備:NanoVue紫外/可見光分光光度計(美國GE Healthcare公司),Agilent 2100 bioanalyzer生物分析儀(美國賽默飛世爾科技公司),熒光顯微鏡(日本Olympus公司),BD FACSCanto流式細胞儀(美國Becton Dickinson公司)。
1.2 細胞培養和分組
培養的BV2細胞分為Man組、Glu組、過表達對照(Con)Glu組(Con Glu組)、過表達TAGLN2 Glu組(TAGLN2 Glu組)、沉默Con Glu組(shCon Glu組)、沉默TAGLN2 Glu組(shTAGLN2 Glu組)。
Man組BV2細胞置于含25 mmol/L Man、25 mmol/L D-Glu、10%胎牛血清的改良Eagle培養基(DMEM培養基)中培養; Glu組、Con Glu組、TAGLN2 Glu組、shCon Glu組、shTAGLN2 Glu組BV2細胞置于含50 mmol/LGlu和10%胎牛血清的DMEM培養基中培養。各組細胞均于37 ℃、5% CO2培養箱中孵育24 h,收集細胞。
1.3 重組慢病毒包裝
根據篩選出沉默效率最高的siRNA序列設計合成短發夾RNA(shRNA)序列,將沉默TAGLN2基因的shRNA序列CCGGGCCGTGAGAACTTCCAGAACTCGAGTTCTGGAAGTTCTCACGGCTTTTT,陰性對照shRNA序列CCGGTTCTCCGAACGTGTCACGTCTCGAGTTCTGGAAGTTCTCACGGCTTTTT分別與慢病毒載體GV248進行連接,并與輔助質粒pHelper1.0、pHelper2.0在293T細胞中包裝成慢病毒GV248-shRNA,感染BV2細胞,4 μg/μl嘌呤霉素篩選,分離單克隆細胞并擴大培養。
合成TAGLN2基因(NM_178598.2)的CDS區序列并與慢病毒載體pHBLV質粒進行連接,與輔助質粒psPAX2、PMD2.G包裝成慢病毒pHBLV-TAGLN2,pHBLV空質粒與輔助質粒包裝成對照慢病毒pHBLV-Con,分別感染BV2細胞,4 μg/μl嘌呤霉素篩選,分離單克隆細胞并擴大培養。
1.4 實時聚合酶鏈反應(RT-PCR)檢測細胞中各因子mRNA相對表達量
采用Trizol試劑提取細胞總RNA,NanoVue紫外/可見光分光光度計檢測RNA樣本濃度,PrimeScript RT Reagent Kit with gDNA Eraser試劑盒將RNA反轉錄成cDNA,使用SYBR Rremix Ex TaqⅡ Kit試劑盒和7500RT-PCR系統進行RT-PCR。每組4個樣本,每個樣本重復3次。相關引物由生工生物工程(上海)股份有限公司合成(表1)。反應條件: 95 ℃預變性30 s,循環1次;95 ℃變性5 s,60℃退火延伸40 s,循環40次;95 ℃變性15 s,60 ℃退火1 min,95 ℃變性15 s,循環1次。以β-actin作為內參照,采用 2?ΔΔCt方法進行結果分析。
1.5 蛋白免疫印跡法檢測細胞中TAGLN2蛋白相對表達量
利用組織蛋白裂解液和蛋白酶抑制劑的混合液提取BV2細胞蛋白,BCA法測定蛋白濃度,取等量蛋白進行十二烷基硫酸鈉聚丙烯酰胺凝膠電泳,并行蛋白質分離且轉印至硝酸纖維素膜上,含10%脫脂牛奶的磷酸鹽緩沖液(PBS)室溫封閉2 h,1∶200稀釋的抗TAGLN2兔多克隆抗體4℃孵育過夜。轉印膜于含0.5%吐溫-20的PBS溶液(PBST)中洗膜10 min,重復3次;1∶3 000稀釋的HRP-IgG室溫孵育2 h,PBST洗膜3次。高敏化學發光試劑盒檢測各組蛋白相對表達量。每組3個樣本,重復1次。
1.6 免疫熒光染色檢測Glu組、Man組BV2細胞中Iba-1表達
4%多聚甲醛固定細胞爬片10 min,0.2% Triton X-100通透30 min,滴加10%山羊血清于室溫下封閉30 min。加入1∶100稀釋的一抗抗CD11b兔多克隆抗體、抗Iba-1羊多克隆抗體,4℃孵育過夜,PBST洗滌3次;加入1∶200稀釋的二抗羊抗兔FITC-IgG、驢抗羊PE-IgG,DAPI避光染色30 min,防熒光淬滅封片劑封片,熒光顯微鏡觀察。細胞中CD11b呈綠色熒光,Iba-1呈紅色熒光,細胞核呈藍色熒光。每組3個樣本,1次免疫熒光染色。
1.7 AimPlex流式高通量多因子檢測BV2細胞中IL-1β、TNF-α、IL-6、干擾素(IFN)-γ、IL-17蛋白表達
收集各組細胞上清液,4 ℃條件下以離心半徑8.6 cm、12 000 r/min離心5 min,25 μl上清液和25 μl樣品稀釋液混勻制成稀釋樣品,標準品稀釋液梯度稀釋標準品。使用Premixed Multiplex Kit Mouse Custom 8-Plex Kit試劑標記標準品或稀釋樣品,依次加入45 μl/孔已包被抗體的混合微球和45 μl/孔的標準品或稀釋樣品,封板,37 ℃、700 r/min避光震蕩60 min。加入1倍洗液100 μl/孔,重復3次;加入1倍生物素標記的二抗25 μl/孔,封板,37℃、700 r/min避光震蕩30 min。加入1倍洗液100 μl/孔,重復2次。加入1倍讀數液200 μl/孔,反復吹吸實驗孔懸浮微球并轉移入5 ml流式細胞管中,使用BD FACSCanto流式細胞儀488 nm激光對反應中的不同大小、不同熒光強度的微球群進行檢測。每組3個樣本,1次檢測。
1.8 轉錄組測序分析
每組設3個生物學重復,Trizol Reagent試劑提取細胞總RNA,NanoDrop超微量分光光度計、Agilent 2100 bioanalyzer生物分析儀檢測RNA濃度、質量,質檢合格構建mRNA文庫,BGI-seq500 平臺進行轉錄組高通量測序,原始測序數據通過SOAPnuke去除低質量、接頭污染以及未知堿基N含量>5%的讀數得到高質量序列。應用HISAT(v2.0.4)、Bowtie2(v2.2.5)分別將高質量序列比對到美國國立生物技術信息中心參考基因組序列GCF_ 000001635. 26_GRCm38.p6、GCF_ 00000 1635.26,得到比對結果。計算每兩個樣品之間所有基因表達量的Pearson相關系數。采用轉錄組學測序技術篩選各組顯著差異表達基因(DEG),篩選標準為|log2[差異表達倍數(FC)]|≥1且P≤0.05[11]。通過RSEM(v1.2.12)軟件計算基因表達水平,DESeq2(v1.4.5)軟件篩選DEG。采用GraphPad Prism軟件繪制火山圖描繪顯著上調基因(log2 FC≥1,P<0.05)和下調基因(log2 FC≤-1,P<0.05)表達的分布情況;R軟件中pheatmap函數繪制聚類熱圖,分析DEG變化;R軟件中的phyper函數進行基因注釋(GO,http://www.geneontology.org/)和京都基因與基因組百科全書(KEGG,https://www.kegg.jp/)富集分析,富集顯著性標準為P<0.05。通過STRING數據庫(https://string-db.org)和Cytoscape軟件進行蛋白-蛋白互作網絡(PPI)分析,圖中節點大小代表該基因權重的高低,節點越大,基因權重越高。
1.9 統計學分析
采用GraphPad Prism軟件進行統計學分析。呈正態分布的計量資料以均數±標準差(±s)表示。組間數據比較采用獨立樣本t檢驗。P<0.05為差異有統計學意義。
2 結果
2.1 高糖誘導小膠質細胞模型建立
高糖誘導小膠質細胞模型建立成功。免疫熒光染色結果顯示,Man組CD11b強表達,Iba-1不表達(圖1A);Glu組CD11b和Iba-1同時強表達(圖1B)。RT-PCR、AimPlex流式高通量多因子檢測結果顯示,與Man組比較,Glu組細胞中IL-1β、TNF-α、IL-6、IFN-γ、IL-17 mRNA(t=-2.877、-7.057、-3.484、-4.603、-6.396)、蛋白表達量(t=-5.598、-5.177、-5.122、-6.178、-7.727)顯著增加,差異有統計學意義(P<0.05)(圖2)。
圖1
甘露醇(Man)組、葡萄糖(Glu)組BV2細胞熒光顯微鏡像(n=3) 1A、1B分別示Man組、Glu組。Iba-1標記的BV2細胞顯示為紅色熒光; CD11b標記的BV2細胞顯示為綠色熒光;4',6-二脒基-2-苯基吲哚標記的細胞核顯示為藍色熒光 標尺:10 μm
圖2
甘露醇(Man)組、葡萄糖(Glu)組BV2細胞中白細胞介素(IL)-1β、腫瘤壞死因子(TNF)-α、IL-6、干擾素(IFN)-γ、IL-17 mRNA、蛋白表達量比較(n=4) 2A示mRNA相對表達量;2B示蛋白相對表達量。*P<0.05,**P<0.01,***P<0.001
2.2 重組慢病毒對BV2細胞中TAGLN2基因的沉默和過表達
與Con Glu組比較,TAGLN2 Glu組細胞中TAGLN2 mRNA、蛋白相對表達量均顯著增加,差異有統計學意義(t=-14.38、-10.43,P<0.001、0.01),過表達TAGLN2的穩轉BV2細胞系構建成功;與shCon Glu組比較,shTAGLN2 Glu組細胞中TAGLN2 mRNA、蛋白相對表達量均顯著降低,差異有統計學意義(t=7.979、4.394,P<0.001)(圖3),沉默TAGLN2的穩轉BV2細胞系構建成功。
圖3
甘露醇(Man)組、葡萄糖(Glu)組、過表達對照(Con)Glu組、過表達TAGLN2 Glu組、沉默Con Glu(shCon Glu)組、沉默TAGLN2 Glu(shTAGLN2 Glu)組細胞中TAGLN2 mRNA和蛋白相對表達量比較(n=3) 3A示mRNA相對表達量,***P<0.001;3B示蛋白相對表達量,*P<0.05,**P<0.01
2.3 DEG表達分析
與Man組比較,Glu組篩選出517個DEG,其中上調、下調基因分別為277、240個(圖4A);與Con Glu組比較,TAGLN2 Glu組共篩選480個DEG,其中上調基因、下調基因分別為147、333個(圖4B); 與shCon組比較,shTAGLN2組共篩選 582個DEG,其中上調、下調基因分別為423、159個(圖4C)。
圖4
差異表達基因分析聚類熱圖 4A示甘露醇(Man)組與葡萄糖(Glu)組;4B示過表達對照(Con)Glu組與過表達轉膠蛋白-2(TAGLN2)Glu組;4C示沉默Con Glu(shCon Glu)組與沉默TAGLN2 Glu(shTAGLN2 Glu)組。紅色越深表示差異表達倍數(FC)越高,綠色越深表示FC越低
2.4 DEG的GO功能富集分析
與Man組相比,Glu組篩選出277個上調DEG顯著富集380條GO條目(P<0.05),主要富集在非經典NF-κB誘導激酶K/NF-κB信號通路及IL-8產生的調控等免疫進程,對氧化應激、炎癥、未折疊蛋白、細胞氧化劑、輻射等刺激應答進程;240個下調DEG顯著富集533條GO條目(P<0.05),主要富集在賴氨酸和蛋白質乙酰化、對轉錄的調控、細胞周期G1/S期轉變的調控、腎小管上皮細胞分化、脂肪細胞分化、橫紋肌細胞分化、神經系統和破骨細胞發育等生物進程。
與Con Glu組相比,TAGLN2 Glu組的147個上調DEG顯著富集392條GO條目(P<0.05),主要富集在脂類、Glu、脂肪酸、甘油醛-3-磷酸生物、三酰甘油、糖原異生和氧化還原等代謝進程;333個下調DEG顯著富集429條GO條目(P<0.05),主要富集在T細胞和B細胞受體信號通路、主要組織相容性復合體(MHC)Ⅱ類分子的抗原加工和提呈、IFN-γ產生等免疫系統進程。
與shCon Glu組相比,shTAGLN2 Glu組的423個上調DEG顯著富集394條GO條目(P<0.05),主要富集在信號素-叢蛋白信號通路、軸突引導中的軸突延伸負調控、鈣離子輸入和轉運、鞘氨醇代謝和脂質磷酸化等進程;159個下調DEG顯著富集333條GO條目(P<0.05),主要富集在對病毒、細菌、IFN-γ、IFN-α和脂多糖應答等先天性免疫應答進程。
GO生物進程分類分析結果顯示,Glu組與Man組相比,免疫系統進程上調基因較下調基因占比增多,代謝進程上調基因較下調基因占比減少(圖5A);TAGLN2 Glu組與Con Glu組相比,免疫系統進程上調基因比下調基因占比減少,代謝進程上調基因較下調基因占比增多(圖5B);shTAGLN2 Glu組與shCon Glu組相比,上調基因和下調基因在免疫系統進程與代謝進程中占比無顯著變化(圖5C)。
圖5
差異表達基因的基因注釋(GO)生物進程分類分析圖 1:細胞進程;2:生物調節;3:生物進程調控;4:代謝進程;5:對刺激的應答;6:多細胞組織進程;7:免疫系統進程;8:發展過程;9:細胞成分組織或生物發生;10:生物進程正調控;11:信號;12:生物進程負調控;13:定位;14:多組織進程 ;15:細胞增生;16:生物進程;17:生殖;18:生殖進程;19:移動;20:生物粘附;21:生長;22:行為;23:解毒作用;24:細胞殺傷;25:節律進程。5A示葡萄糖(Glu)組與甘露醇(Man)組;5B示過表達轉膠蛋白-2(TAGLN2)Glu組與過表達對照(Con)Glu組;5C示沉默TAGLN2 Glu(shTAGLN2 Glu)組與沉默Con Glu(shCon Glu)組。紅色條柱代表上調基因富集的GO條目基因數占比,藍色條柱代表下調基因富集的GO條目基因數占比
2.5 DEG的KEGG通路富集分析
與Man組相比,Glu組上調DEG顯著富集16條KEGG條目(P<0.05),主要與NF-κB、Jak-信號轉導和轉錄激活因子(STAT)、 IL-17信號通路等免疫系統進程相關;下調DEG顯著富集17條KEGG條目(P<0.05),主要與糖胺多聚糖、賴氨酸、膽固醇等代謝進程相關(圖6)。
圖6
葡萄糖組與甘露醇組差異表達基因生物進程顯著富集程度前20的京都基因與基因組百科全書條目1:核轉錄因子-κB信號通路;2:蛋白酪氨酸激酶- 信號傳導及轉錄激活蛋白信號通路;3:Toll樣受體信號通路;4:白細胞介素-17信號通路;5:腫瘤壞死因子信號通路;6:趨化因子信號通路;7:輔助性T 細胞17細胞分化;8:核苷酸寡聚化結構域樣受體信號通路;9:氧化磷酸化;10:C5-支二元酸代謝;11:谷胱甘肽代謝;12:逆行內源性大麻素信號;13:生熱作用;14:心肌收縮;15:磷脂酰肌醇三羥基激酶-蛋白激酶B信號通路;16:核糖體;17:線粒體自噬;18:粘附連接;19:低氧誘導因子-1信號通路;20:腎素-血管緊張素系統相關蛋白1信號通路;21:Wnt信號通路;22:Notch信號通路;23:長時程增強;24:黑素原生成;25:甲狀腺激素信號通路;26:甘油脂代謝;27:甲狀旁腺激素的合成和分泌;28:檸檬烯和蒎烯的降解;29:昆蟲激素的生物合成;30:膽固醇代謝;31:賴氨酸降解;32:醛固酮的合成和分泌;33:糖胺多聚糖的降解。橫軸左、右側柱狀圖分別代表上調基因、下調基因,折線圖代表該條目包含的基因數
與Con Glu組相比,TAGLN2 Glu組上調DEG顯著富集44條KEGG條目(P<0.05),主要與碳、脂肪酸、甘油脂、丙酮酸、氯代烷和氯烯烴、氨基酸等代謝進程相關;下調DEG顯著富集25條KEGG條目(P<0.05),主要與T細胞與B細胞受體信號通路,Th1、Th2、Th17細胞分化,腸道免疫網絡與IgA產生,NF-κB、TNF、Jak-STAT、趨化因子信號通路以及白細胞、跨內皮細胞遷移等免疫系統進程相關(圖7)。
圖7
過表達轉膠蛋白-2葡萄糖組與過表達對照葡萄糖組差異表達基因生物進程顯著富集程度前20的京都基因與基因組百科全書條目 1:不同環境中的微生物代謝;2:代謝通路;3:碳代謝;4:脂肪酸降解;5:脂肪酸代謝;6:甘油脂代謝;7:丙酮酸代謝;8:氯代烷和氯烯烴的降解;9:糖酵解/糖異生;10:纈氨酸、亮氨酸和異亮氨酸的降解;11:氨基酸的生物合成;12:次生代謝物的生物合成;13:抗生素的生物合成;14:生熱作用;15:胰島素信號通路;1:6:脂肪細胞脂解作用的調控;17:近端小管碳酸氫鹽回收;18:過氧化物酶體增生物激活受體信號通路;19:腺苷5'-單磷酸激活蛋白激酶信號通路;20過氧物酶體;21:緊密連接;22:灶性粘連;23:腎素-血管緊張素系統信號通路;24:細胞粘附分子;25:長時程增強;26:血小板活化;27:鞘糖脂的生物合成;28:趨化因子信號通路;29: Fc段γ受體介導的吞噬作用;30:蛋白酪氨酸激酶- 信號傳導及轉錄激活蛋白信號通路信號通路;31:原處理和提呈;32:腫瘤壞死因子信號通路;33:白細胞跨內皮細胞遷移;34:核因子-κB信號通路;35:腸道免疫網絡與免疫球蛋白A的產生;36:輔助性T細胞(Th)17細胞分化;37:B細胞受體信號通路;38:Th1和Th2細胞分化;39:T細胞受體信號通路;40:造血細胞譜系。橫軸左、右側柱狀圖分別代表上調基因、下調基因,折線圖代表該條目包含的基因數
與shCon Glu組相比,shTAGLN2 Glu組上調DEG顯著富集40條KEGG條目(P<0.05),主要與TNF、趨化因子信號通路等免疫進程及色氨酸、環氧氯己烷、鞘脂、鞘糖脂和丁酸等代謝進程相關;下調DEG顯著富集14條KEGG條目(P<0.05),主要與核苷酸寡聚化結構域樣受體、Toll樣受體信號通路以及氮代謝、脂肪消化吸收等進程相關(圖8)。
圖8
沉默轉膠蛋白-2葡萄糖組與沉默對照葡萄糖組差異表達基因生物進程顯著富集程度前20的京都基因與基因組百科全書條目 1:腫瘤壞死因子信號通路;2:趨化因子信號通路;3:輔助性T細胞(Th)1和Th2細胞分化;4:細胞因子-細胞因子受體相互作用;5:色氨酸代謝;6:環氧氯己烷與氯苯的降解;7:氟苯甲酸鹽的降解;8:鞘脂代謝;9:糖脂的生物合成;10:甲苯降解;11:丁酸代謝;12:軸突引導;13:醛固酮的合成和分泌;14:膽堿能突觸;15:催產素信號通路;16:多巴胺能突觸;17:鈣信號通路;18:心血管活性多肽信號通路;19:環磷酸鳥苷-蛋白激酶G信號通路;20:絲裂原活化蛋白激酶信號通路;21:肌動蛋白細胞骨架的調控;22:膽堿能突觸;23:松弛素信號通路;24:胞質DNA傳感途徑;25:破骨細胞分化;26:內分泌和其他因素調節的鈣重吸收;27:軸突引導;28:脂肪消化吸收;29:細胞色素P450對外源物質的代謝作用;30:纈氨酸、亮氨酸和異亮氨酸的生物合成;31:氮代謝;32:Toll樣受體信號通路;33:視黃酸(維甲酸)誘導基因蛋白-I樣受體信號通路;34:核苷酸寡聚化結構域樣受體信號通路。橫軸左、右側柱狀圖分別代表上調基因、下調基因,折線圖代表該條目包含的基因數
2.6 KEGG免疫系統進程和免疫相關DEG的PPI分析
與Man組相比,Glu組有58個與免疫相關DEG,8條免疫系統進程。包括Stat1、Stat2和Mef2c 3個轉錄因子,權重最高的10個DEG分別為Il1b、Stat1、Tlr4、Cxcl2、Tlr7、Cd14、Stat2、Csf1、Cxcr4、Tlr9(圖9A)。
圖9
蛋白-蛋白互作網絡圖 NF:核因子;Jak-STAT:蛋白酪氨酸激酶- 信號傳導及轉錄激活蛋白;IL-白細胞介素;Th:輔助性T細胞;NOD:核苷酸寡聚化結構域;FcγR:Fc段γ受體;IgA:免疫球蛋白A;TNF:腫瘤壞死因子;RIG:視黃酸(維甲酸)誘導基因蛋白;9A示葡萄糖(Glu)組與甘露醇(Man)組;9B示過表達轉膠蛋白-2(TAGLN2)Glu組與過表達對照(Con)Glu組;7C示沉默TAGLN2 Glu(shTAGLN2 Glu)組與沉默Con Glu(shCon Glu)組。紅色越深表示差異基因表達量越高,綠色越深表示差異基因表達量越低;藍色、紫色越深表示P值越大
與Con Glu組相比,TAGLN2 Glu組有104個與免疫相關DEG,13條免疫系統進程。包括Stat4、Zbtb1 2個轉錄因子,權重最高的10個DEG分別是Ptprc、Lck、Cd19、Vav1、Cd3g、Ctla4、Cd5、Cd3e、Itk、Cd3d(圖9B)。
與shCon Glu組相比,shTAGLN2 Glu組有68個與免疫相關DEG,7條免疫系統進程,包括Fos、Pou2f2、Maf、Mef2c和Irf7等5個轉錄因子,權重最高的10個DEG分別是Cxcl10、Irf7、Tlr3、Ifih1、Ifi204、Ifit1、Ifit3、Dhx58、Cd274、Trim30a(圖9C)。
2.7 TAGLN2抑制高糖誘導的炎癥因子表達
RT-PCR檢測結果顯示,與Man組比較,Glu組細胞中Card11、Icos、Chst3、Kynu mRNA相對表達量顯著增高,差異有統計學意義(t=-5.925、-6.339、-5.523、-5.948,P=0.001、0.001、0.003、0.001)。與Con Glu組比較,TAGLN2 Glu組中Card11、Icos、Chst3、Kynu mRNA相對表達量顯著降低,差異有統計學意義(t=13.530、3.482、6.949、5.399,P=0.000、0.018、0.001、0.006)(圖10A)。
圖10
甘露醇(Man)組、葡萄糖(Glu)組、過表達對照(Con)Glu組、過表達TAGLN2 Glu組、沉默Con Glu(shCon Glu)組、沉默TAGLN2 Glu(shTAGLN2 Glu)組細胞中各因子mRNA和蛋白相對表達量比較(n=3) 10A示Card11、Icos、Chst3、Kynu mRNA相對表達量,*P<0.05,**P<0.01,***P<0.001;10B示白細胞介素(IL)-1β、腫瘤壞死因子(TNF)-α、IL-6、干擾素(IFN)-γ、IL-17 mRNA相對表達量,*P<0.05,**P<0.01;10C示IL-1β、TNF-α、IL-6、IFN-γ、IL-17 蛋白相對表達量,*P<0.05,**P<0.01
RT-PCR、AimPlex流式高通量多因子檢測結果顯示,與Con Glu組比較,TAGLN2 Glu組細胞中IL-1β、TNF-α、IL-6、IFN-γ、IL-17 mRNA(t=2.960、5.800、3.130、7.690、6.530)、蛋白相對表達量(t=3.050、4.530、3.420、6.880、3.380)均顯著下降,差異有統計學意義(P<0.05)(圖10B,10C)。
3 討論
既往研究表明,高糖能誘導小膠質細胞NF-κB信號通路激活[3-4],也能誘導視網膜血管內皮細胞和視網膜中Jak-STAT信號通路活化,并釋放大量炎癥因子和趨化因子[12-13]。本研究結果顯示,高糖能夠激活BV2細胞中NF-κB、Jak-STAT信號通路,促進TNF-α、IL-6和IL-1β等多種炎癥因子和趨化因子釋放。研究表明,TAGLN2過表達抑制淋巴毒素-β受體激動劑誘導的血管平滑肌細胞中NF-κB誘導激酶及其下游NF-κB的典型和非典型信號通路的表達[14]。本研究也發現過表達TAGLN2能夠顯著抑制高糖誘導的NF-κB信號通路的激活和炎癥因子、趨化因子的釋放。GO富集分析也發現過表達TAGLN2后小膠質細胞中T細胞和B細胞受體信號通路、MHC Ⅱ類分子的抗原加工和提呈、白細胞與細胞粘附及調節和IFN-γ產生的正向調節等免疫系統進程被顯著抑制。沙門菌感染后引起TAGLN2?/?小鼠的血漿TNF-α、IL-6、IL-1β、IL-12水平顯著升高[9]。本研究結果顯示,沉默TAGLN2后高糖誘導的小膠質細胞中TNF信號通路、趨化因子和細胞因子-細胞因子受體相互作用信號通路被激活,TNF-α、IL-6、IL-17、IL-1β等炎癥因子和趨化因子表達增加,但是Toll樣受體信號通路和視黃酸(維甲酸)誘導基因蛋白-Ⅰ樣受體信號通路等模式識別受體信號通路被抑制;GO富集分析也發現沉默TAGLN2后小膠質細胞對病毒、細菌、IFN-α、IFN-β和脂多糖的固有免疫應答進程減少,表明沉默TAGLN2后小膠質細胞識別抗原和外部刺激能力減弱,炎癥反應增加,免疫功能失衡。因此,TAGLN2在高糖誘導的小膠質細胞炎癥反應中發揮了關鍵作用,是治療DR的潛在藥物靶點。
本研究結果顯示,分別與Man組、Con Glu組、shCon Glu組這3個對照組比較,Glu組、TAGLN2 Glu組及shTAGLN2 Glu組同時表達的DEG為14個,其中Card11、Icos、Chst3、Kynu均與疾病炎癥反應相關。Card11基因參與T細胞受體、B細胞受體介導的NF-κB信號通路活化[15-16]。Icos基因介導非肥胖糖尿病(NOD)小鼠胰島素依賴型糖尿病發展,是NOD小鼠發生胰島素炎和高血糖所不可缺少的;其缺失可導致Th1細胞因子IFN-γ的產生減少[17-18]。抑制Icos/Icos配體信號顯著抑制炎癥因子的表達,以及巨噬細胞、中性和嗜酸性粒細胞浸潤[19-20]。Chst3基因參與硫酸軟骨素生物合成和糖胺聚糖代謝,其沉默可減少肺氣腫巨噬細胞的積累并抑制基質金屬蛋白酶-9的表達,促進疾病恢復[21]。Kynu是牛皮癬和其他炎性疾病中的新型炎性因子[22],抑制小膠質細胞中的Kynu通路能夠保護反應性小膠質細胞相關神經元的萎縮[23]。本研究結果顯示,過表達TAGLN2能夠顯著抑制高糖誘導的小膠質細胞中Card11、Icos、Chst3、Kynu的表達,減少NF-κB信號通路活化和多種炎癥因子和趨化因子的表達。
本研究存在的不足:(1)DR發病機制復雜,高血糖僅是疾病發生的主要原因之一,高糖干預的細胞模型不能完全模擬體內情況,還需要進一步在體實驗加以驗證;(2)本研究僅檢測了高糖處理24 h顯著DEG及信號通路的改變,其中關鍵信號通路的功能分析驗證及更長時間高糖處理DEG和信號通路的改變,仍需要進一步實驗研究。