海馬和前額葉皮層之間神經信息的傳遞與交互在學習和記憶功能的實現中發揮了重要作用,但記憶相關任務學習前后,兩腦區之間的連接特性是否及如何變化,仍有待深入研究。本文應用在體微電極記錄技術,獲取了任務學習前后,8只大鼠執行T迷宮記憶任務時腹側海馬(vHPC)和內側前額葉皮層(mPFC)的局部場電位(LFPs)信號,并基于動態因果模型(DCM)對比分析大鼠學習前后執行T迷宮記憶任務時vHPC和mPFC之間的因果連接特性的變化。結果顯示,大鼠執行T迷宮記憶任務時,從vHPC到mPFC為前向連接,從mPFC到vHPC為后向連接。任務學習后兩腦區之間的前向連接增強,后向連接減弱,與大鼠正確執行任務時兩腦區連接變化趨勢一致。綜上所述,通過本文研究,希望能夠為進一步從不同腦區連接特性變化角度研究學習記憶的相關機制提供支持。
引用本文: 李雙燕, 鄭衛然, 阿蘭, 王龍龍, 劉素紅, 劉慧. 學習對記憶任務中大鼠海馬—前額葉連接特性影響的研究. 生物醫學工程學雜志, 2024, 41(6): 1095-1102. doi: 10.7507/1001-5515.202312042 復制
版權信息: ?四川大學華西醫院華西期刊社《生物醫學工程學雜志》版權所有,未經授權不得轉載、改編
0 引言
學習與記憶是通過獲取、儲存和更新外部信息來實現認知活動和自我提升的關鍵[1]。學習與記憶作為大腦的高級認知活動,往往需要多個腦區協同參與才能實現[2-3]。其中,海馬(hippocampus,HPC)和前額葉皮層(prefrontal cortex,PFC)之間神經元集群活動的協同和神經信息的交互,被認為在學習與記憶功能的實現中發揮了重要作用[4-7]。例如,Morici等[8]研究發現,在對過去事件進行回憶的過程中,大鼠PFC和腹側HPC(ventral HPC,vHPC)之間的局部場電位(local field potentials,LFPs)信號同步性顯著增加。而PFC和HPC腦區之間功能連接的異常,可能是造成唐氏綜合征患者記憶功能障礙的神經機制之一[9]。此外,Avigan等[10]在大鼠十字迷宮任務的學習過程中,通過選擇性地抑制PFC或HPC中神經元的活性,發現HPC的雙側失活,嚴重損害了大鼠的空間學習和記憶功能,而雙側PFC的失活主要損害了大鼠的辨別學習能力,表明空間學習功能的實現與PFC和HPC之間的相互作用密切相關。雖然這些研究表明,HPC和PFC之間神經信息的傳遞和相互作用對于實現學習與記憶功能具有關鍵作用,但記憶相關任務學習前后,兩腦區之間的連接特性是否及如何變化,仍有待深入研究。
動態因果模型(dynamic causal modeling,DCM)是由Friston等[11]提出的一種從有效連通性角度探索腦功能相關問題的方法,目前已經在神經科學領域得到了廣泛應用[12-17]。與其它分析腦區間神經信息相互作用的方法相比,DCM模型能夠將外界環境(輸入刺激)、神經元集群的活動狀態,以及記錄所得神經電活動信號特征三者相聯立,更具有生物物理合理性[18-20]。因此,本文擬應用符合神經生理學特性的DCM算法,探究大鼠學習前后執行記憶任務過程中vHPC和內側PFC(medial PFC,mPFC)之間的因果連接特性的變化。
綜上,本文應用在體微電極記錄技術,獲取了8只大鼠在T迷宮記憶任務學習前后執行該任務過程中vHPC和mPFC腦區的LFPs信號。隨后,基于DCM模型構建了兩腦區之間可能的連接模型,并應用貝葉斯算法進行了最優模型選擇。最后,基于最優調制模型,對比分析了大鼠在T迷宮記憶任務學習前后,正確與錯誤執行該任務時vHPC和mPFC之間連接強度的變化。通過開展上述研究,本文期望能夠從腦區間因果連接特性變化的角度探索腦區間神經信息交互處理的相關機制,為今后相關領域研究奠定理論和實踐基礎。
1 材料與方法
實驗對象為8只成年雄性維斯塔爾(Wistar)大鼠,體重280~320 g,由北京華阜康生物科技股份有限公司提供。飼養期間,不限制大鼠對水和食物的獲取。任務訓練開始后,實驗動物可以獲得能夠維持其體重不低于正常體重85%的食物。本研究中的所有實驗流程與環節均嚴格遵循科研倫理規范,實驗獲河北工業大學生物醫學倫理委員會批準(審查編號:HEBUTaCUC2022062)。
1.1 T迷宮記憶任務及LFPs數據獲取
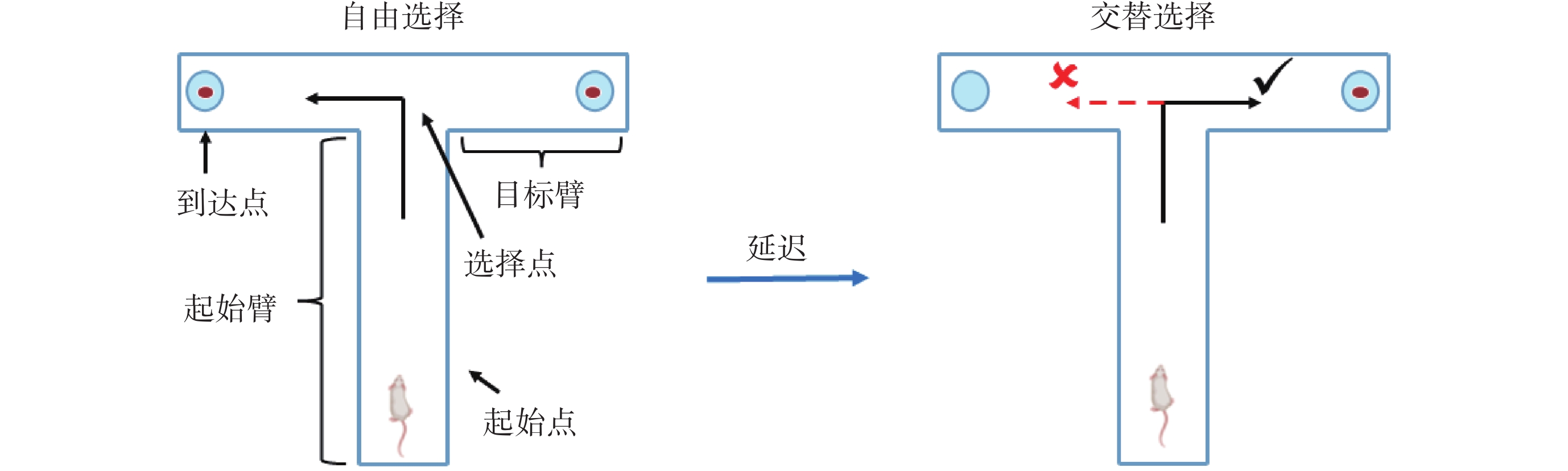
實驗開始前,需先將微電極植入大鼠mPFC和vHPC腦區。具體過程見文獻[21],簡述如下:將麻醉中的大鼠頭部固定到立體定位儀上,并使其顱骨暴露。在非目標腦區上方的顱骨固定3~4顆螺釘作為固定和接地電極。隨后,去除mPFC(前囟前2.50~4.50 mm,旁開0.20~1.00 mm)和vHPC(前囟后4.60~6.00 mm,旁開4.40~5.40 mm)上方的顱骨及硬腦膜。最后,使用微推進器P_PAD LITE(Scientifica Inc.,美國)將鎳鉻合金電極ST35(Plexon Inc.,美國)推進至目標腦區對應深度(mPFC:2.50~3.50 mm,vHPC:7.00~8.00 mm)后,使用牙科水泥進行固定。手術結束后,待大鼠恢復5~7 d,開始進行T迷宮記憶任務[22]學習并同步采集神經電生理信號。如圖1所示,T迷宮各臂寬15 cm,高20 cm,起始臂長70 cm,兩個目標臂的長度各為50 cm。T迷宮記憶任務主要包括自由選擇、延遲、交替選擇三個階段。在自由選擇階段,大鼠選擇任意目標臂均可獲得食物獎勵。隨后,經過起始點5~10 s的延遲等待后,大鼠進入交替選擇階段。在該階段,大鼠需選擇與前面自由選擇階段相異的目標臂,才可以再次得到食物獎勵,這被視為任務正確執行的標準。反之,視為錯誤執行任務。在大鼠執行任務的正確率連續2 d達到80%及以上之后,任務學習階段結束。
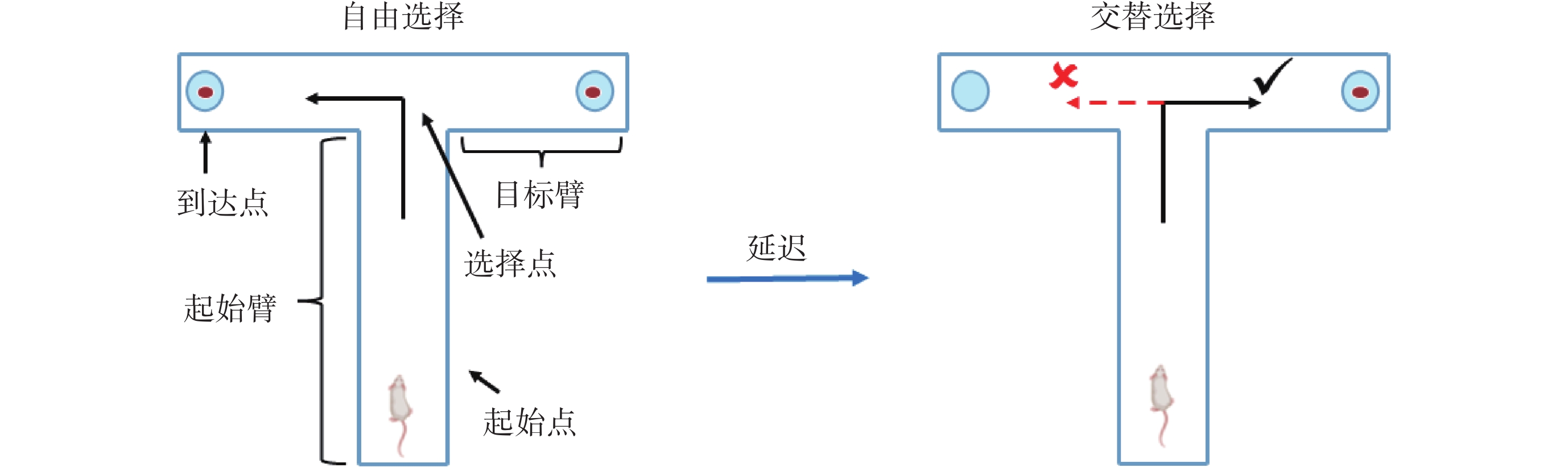 圖1
T迷宮記憶任務示意圖
Figure1.
Diagram of the T-maze memory task
圖1
T迷宮記憶任務示意圖
Figure1.
Diagram of the T-maze memory task
在大鼠執行T迷宮記憶任務時應用神經電生理信號采集系統OmniPlex/128(Plexon Inc.,美國)同步記錄獲取大鼠vHPC和mPFC腦區的LFPs信號(采樣頻率1 000 Hz)。在應用DCM模型對LFPs信號進行分析之前,首先對其進行4~100 Hz的濾波以及陷波濾波,以消除50 Hz工頻干擾。
為探索學習前后大鼠在執行T迷宮記憶任務過程中,其vHPC和mPFC之間連接特性的變化,本文在后續研究中,對應任務學習前后狀態,分別選用了任務學習初始階段(開始任務訓練后第1~3天)和學習階段結束后(開始任務訓練后第8~12天),大鼠在執行任務過程中的LFPs數據進行DCM模型分析。其中,包括任務學習前:8只大鼠,共96次實驗;任務學習后:8只大鼠,共111次實驗。
1.2 DCM模型
DCM模型基于動態系統建模和貝葉斯統計框架,對神經網絡中的有效連接模式及其調節機制進行研究。該模型將大腦抽象為一個復雜的“刺激輸入—內部狀態變化—輸出響應”的動態系統,其中,外部環境的刺激作為輸入,能夠激發神經系統內部狀態的動態演變,而這些內部狀態的變化可以通過特定的輸出信號得以體現。在這一理論架構下,DCM模型巧妙利用了給定的刺激輸入與相應的輸出反饋數據,通過調整模型的參數,逆向推斷出隱藏于特定腦區背后的神經元活動變化及其相互間的有效連接模式。DCM模型中用于描述外界刺激下神經元狀態變化的狀態方程,其表達式如式(1)所示:
 |
其中,?表示神經元狀態變量;F是描述外部輸入刺激u或神經元活動z對其他腦區神經元狀態變化產生生理影響的非線性函數;θ表示激活腦區生成模型中一系列待定的生物物理參數和連接參數;該方程反映了不同神經元集群之間的信息傳遞過程[23]。
此外,基于觀測方程,可以實現從目標腦區神經元集群活動狀態到測量所得信號的進一步映射[24],其表達式如式(2)所示:
 |
其中,觀測信號y通過觀測函數g與隱含狀態z及觀測參數?之間建立了明確的依賴關系。
在運用DCM模型解析神經元集群間有效連接模式的過程中,確保模型的神經生物學合理性是保障分析結果準確性的前提。這要求模型參數必須富含生物物理學信息,從而增加了參數估計的復雜性與挑戰性[25]。為了高效且可靠地實現對模型參數的估計,DCM模型采用了基于貝葉斯統計學的反演方法[11]。
根據貝葉斯原理,對于特定的DCM模型m,其參數 的后驗概率密度p(θ|y, m)的計算,如式(3)所示:
的后驗概率密度p(θ|y, m)的計算,如式(3)所示:
 |
其中,p(y|θ, m)表示觀測數據y在給定參數θ與模型m條件下的似然性;p(θ|m)作為參數的先驗分布,融入了研究者對未知參數的先驗知識;而p(y|m),即模型證據值或邊際似然函數,則綜合評估了模型m對數據 的整體解釋能力,其在模型選擇與評估中扮演著至關重要的角色[26]。
的整體解釋能力,其在模型選擇與評估中扮演著至關重要的角色[26]。
基于模型證據值,可以對相同樣本數據(如多腦區神經信號)所可能具有的不同連接模型的合理性進行比較[11]。模型間的比較由貝葉斯因子體現,表達式如式(4)所示:
 |
其中,mi和mj分別代表兩種腦區間連接方式不同的DCM模型。貝葉斯因子BFi, j用于量化比較這兩個模型與觀測數據之間的擬合程度。具體而言,當BFi, j>1時,這指示了相較于模型mj,模型mi在解釋觀測數據方面展現出更高的證據支持,即模型mi與觀測數據的匹配性更為優越[26]。為解決由于多個貝葉斯因子相差過大而產生的難以表示的問題,可進一步對上式兩邊取對數,應用相對對數模型證據值進行表示,如式(5)所示:
 |
根據相關文獻[27-28],本研究中假設同側mPFC和vHPC之間存在雙向連接。隨后,應用貝葉斯反演和模型的相對對數證據值進行最優連接模型選擇,并確定學習這一影響因素最可能對腦區間連接產生影響的情況,即最優調制模型;其中,相對對數模型證據值最大的模型,即為最優連接模型或最優調制模型。同時,基于模型后驗概率評估所選最優連接模型和調制模型的置信度,當后驗概率越接近1時,所選最優連接模型和最優調制模型的置信度越高[19]。應用上述方法,本文對比分析了大鼠學習T迷宮記憶任務前后,以及正確和錯誤執行任務時,mPFC和vHPC之間連接強度的變化,探索了學習對相關腦區神經信息傳遞的影響。本文中的DCM模型分析使用英國倫敦大學學院開發的腦成像數據序列處理軟件Statistical Parametric Mapping 12進行。
2 結果
2.1 vHPC和mPFC間最優連接模型選擇結果
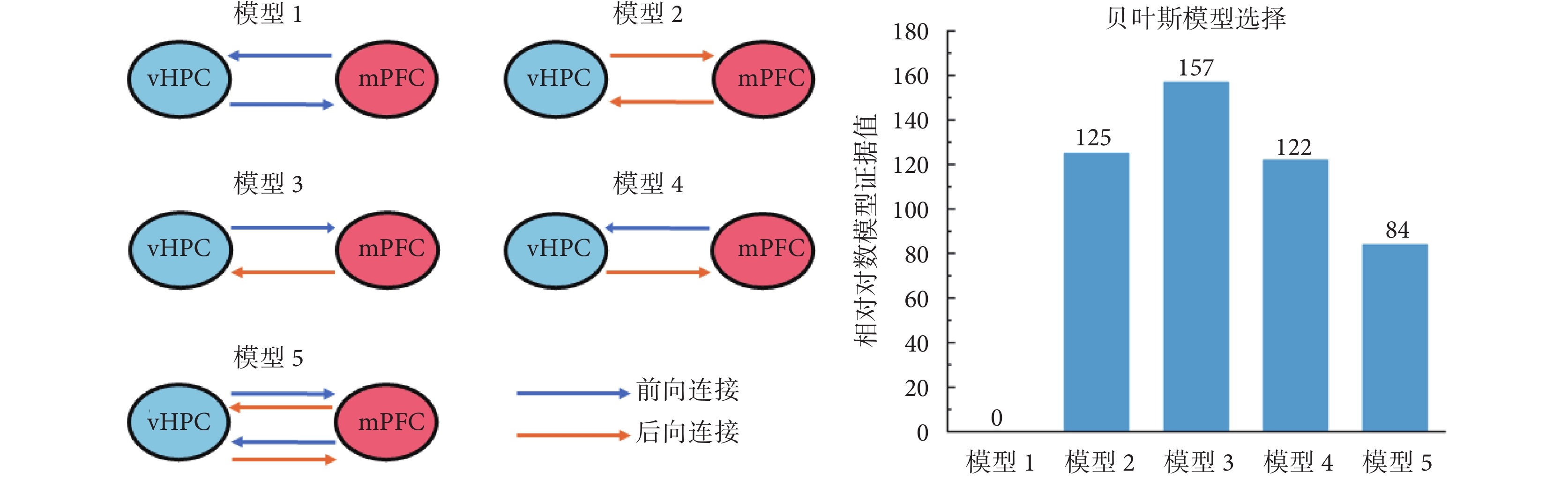
如圖2所示,為vHPC和mPFC之間的連接模型及貝葉斯模型選擇結果。根據相關文獻[27],同側mPFC和vHPC之間可能存在5種不同情況的雙向連接,分別為:模型1,兩腦區之間存在雙向前向連接;模型2,兩腦區之間存在雙向后向連接;模型3,vHPC到mPFC為前向連接,mPFC到vHPC為后向連接;模型4,與前一模型相反,mPFC到vHPC為前向連接,vHPC到mPFC為后向連接;模型5,兩腦區之間同時存在雙向前向連接和雙向后向連接。本研究中,首先基于8只大鼠學習前和學習后執行T迷宮記憶任務時的LFPs數據,應用貝葉斯反演和模型的相對對數證據值進行上述模型的最優選擇。貝葉斯模型選擇結果顯示,模型3具有最大的相對對數模型證據值(157),即vHPC和mPFC之間最優的連接模型為模型3(vHPC到mPFC為前向連接,mPFC到vHPC為后向連接)。進一步對該模型的置信度進行分析的結果顯示,其后驗概率為1,模型可信。因此,接下來,進一步基于模型3,對大鼠學習前后連接受影響的可能情況進行分析。
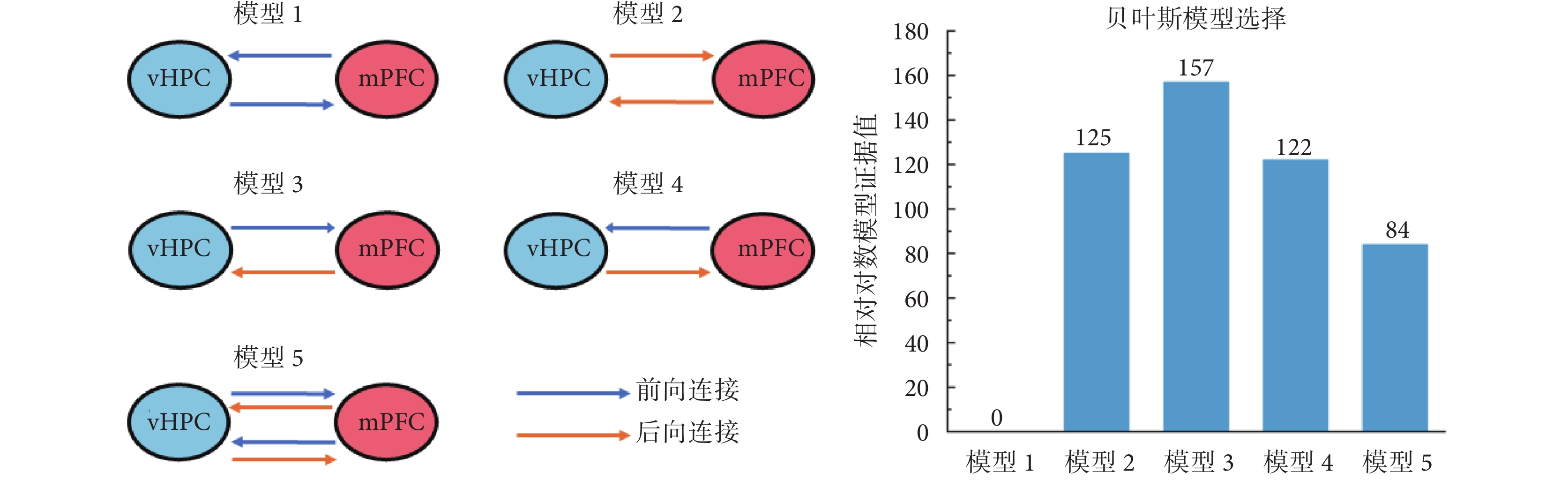 圖2
5種源間連接類型的描述和貝葉斯模型比較結果
Figure2.
Illustration of 5 different between-source connection models and Bayesian model comparison results
圖2
5種源間連接類型的描述和貝葉斯模型比較結果
Figure2.
Illustration of 5 different between-source connection models and Bayesian model comparison results
2.2 最優調制模型選擇結果
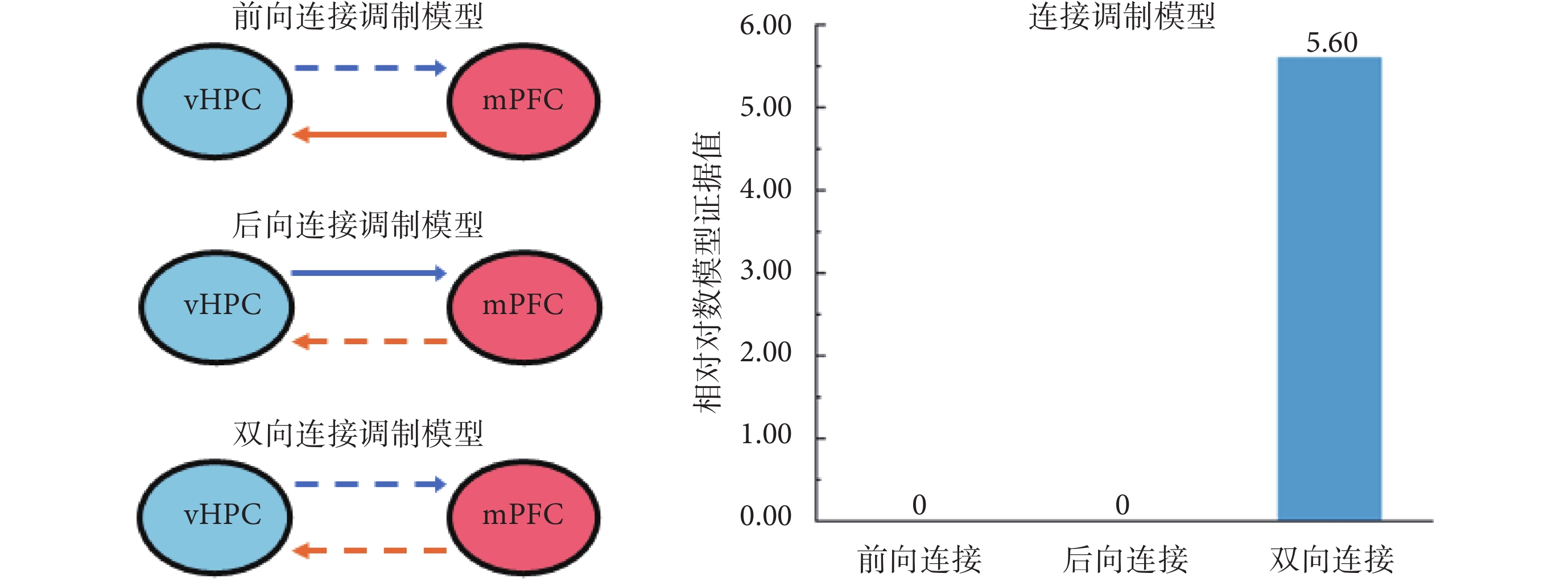
基于所選擇的最優連接模型,學習這一因素對vHPC和mPFC之間連接的影響可能存在以下3種情況:一是僅vHPC到mPFC的前向連接受到影響;二是僅mPFC到vHPC的后向連接受到影響;三是兩腦區之間的雙向連接,即前向和后向連接均受到了影響。因此,所構建的連接調制模型如圖3所示,其中,虛線表示受外界因素調制的腦區間連接,實線表示未受到影響的連接。同樣,應用貝葉斯反演和模型的相對對數證據值對上述模型進行最優選擇。圖3右圖連接調制模型中,橫軸分別對應前向連接受影響、后向連接受影響、雙向連接受影響的調制模型分析結果,縱軸為相對對數模型證據值。從圖3中可以看出,雙向連接均受到影響的模型具有最大的相對對數模型證據值(5.60),是最優的調制模型。進一步對其置信度進行分析的結果表明,該模型的后驗概率為0.98,具有較高置信度(98%)。上述結果表明,對記憶任務的學習能夠同時影響vHPC和mPFC之間的雙向連接。
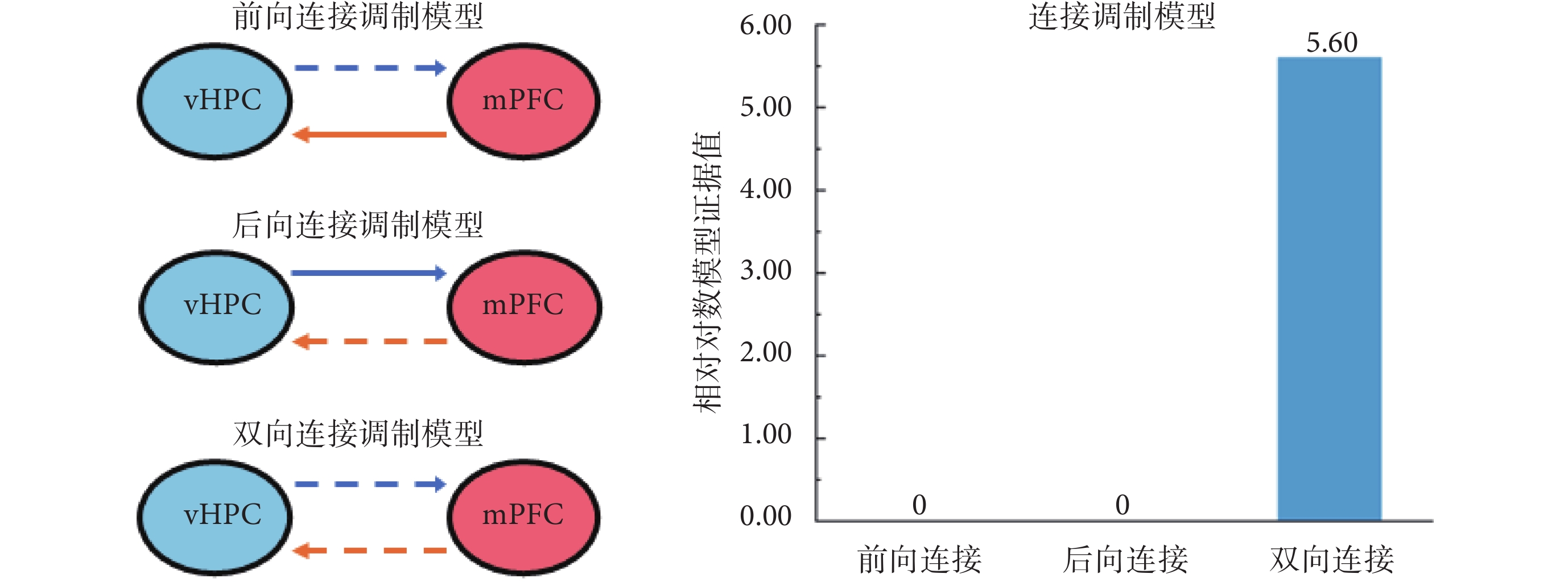 圖3
3種連接調制模型的描述和貝葉斯模型比較結果
Figure3.
Illustration of 3 different connection modulation models and Bayesian model comparison results
圖3
3種連接調制模型的描述和貝葉斯模型比較結果
Figure3.
Illustration of 3 different connection modulation models and Bayesian model comparison results
2.3 學習前后vHPC和mPFC之間雙向連接強度的變化
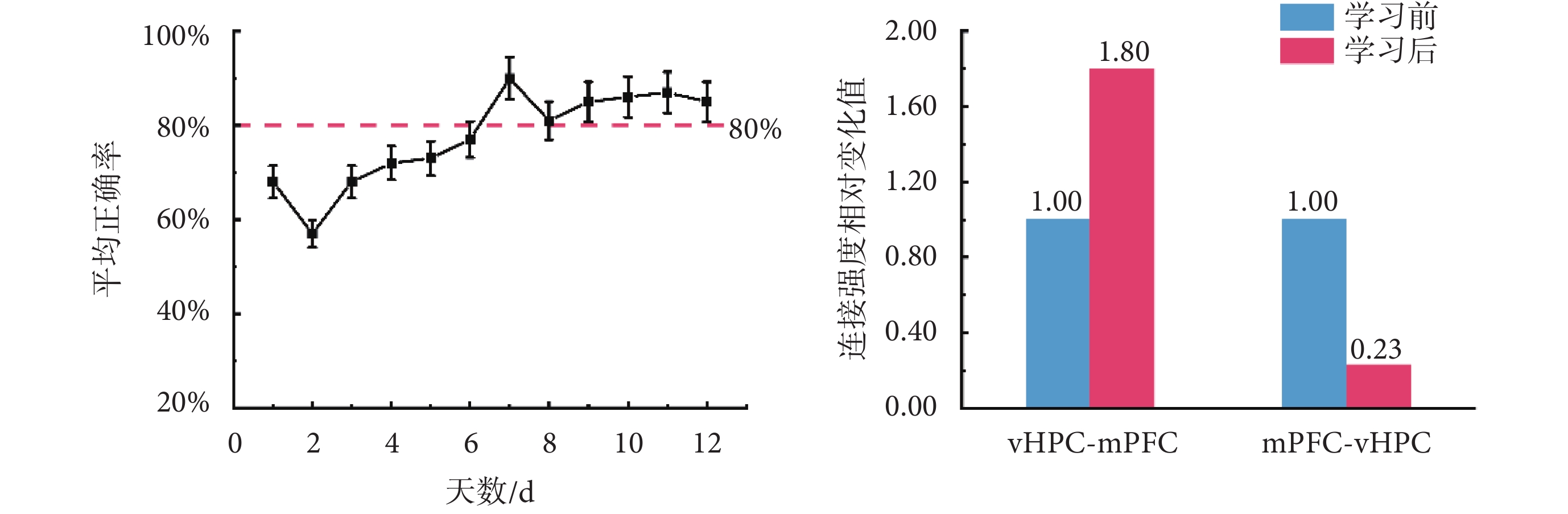
為研究學習前后vHPC和mPFC之間的雙向連接如何變化,本研究進一步基于任務學習前后,大鼠在執行T迷宮記憶任務過程中兩腦區的LFPs信號,應用所選最優調制模型,對任務學習前后兩腦區之間連接強度的變化進行了分析,結果如圖4所示。其中,8只大鼠每天的T迷宮記憶任務平均正確率(以均值±標準差表示)結果顯示,隨著訓練天數的增加,大鼠執行T迷宮記憶任務的平均正確率總體呈上升趨勢,最終能夠達到并維持在80%以上。如前文所述,本文中選取了每只大鼠任務學習初始階段(第1~3天)和學習階段結束后(第8~12天)的神經電活動信號進行了分析。本文基于學習前后大鼠執行T迷宮記憶任務時vHPC和mPFC的LFPs信號,應用DCM模型,對兩腦區之間連接強度變化進行了分析。圖4右圖橫坐標軸上,vHPC-mPFC,表示從vHPC到mPFC的前向連接強度在學習前后的對比結果;mPFC-vHPC,表示從mPFC到vHPC的后向連接強度在學習前后的對比結果。圖4右圖縱坐標軸,表示連接強度相對變化值。
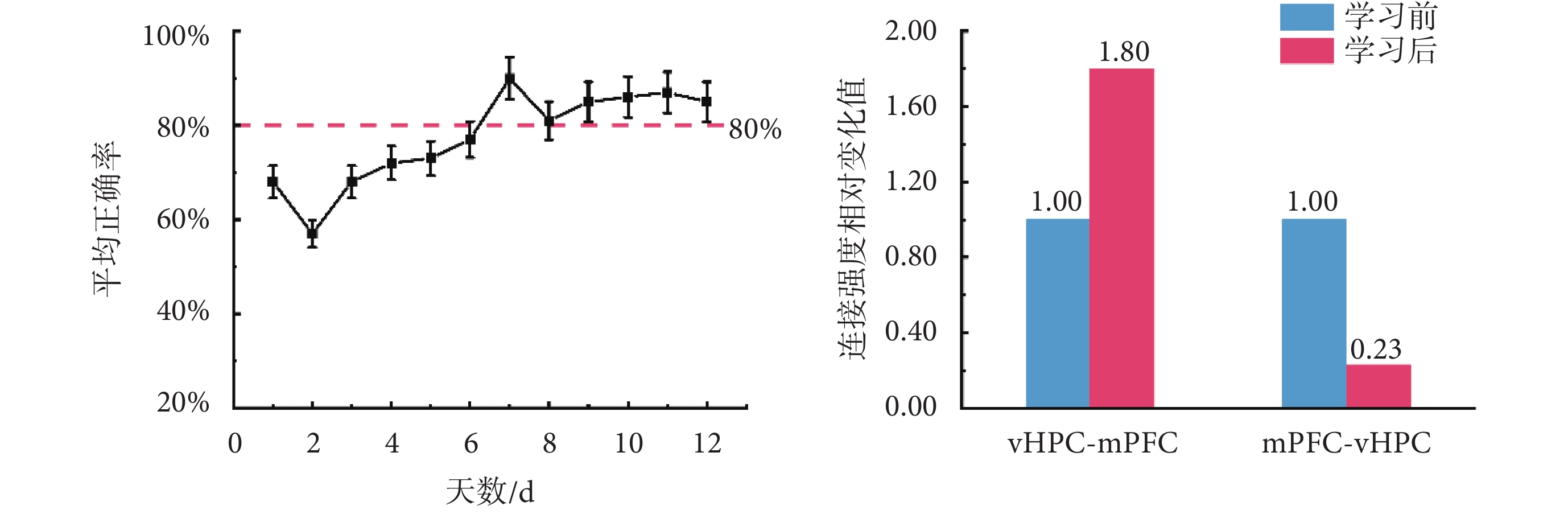 圖4
大鼠學習 T 迷宮任務并比較 vHPC 和 mPFC 之間的連接強度
Figure4.
Changes in T maze task accuracy during rats learning and a comparison of connection strength between the vHPC and mPFC pre- and post-learning
圖4
大鼠學習 T 迷宮任務并比較 vHPC 和 mPFC 之間的連接強度
Figure4.
Changes in T maze task accuracy during rats learning and a comparison of connection strength between the vHPC and mPFC pre- and post-learning
結果顯示,相比學習前,學習后大鼠在執行任務時,vHPC和mPFC兩腦區之間的前向連接(vHPC-mPFC)增強為學習前的1.80倍,而后向連接(mPFC-vHPC)降低為0.23倍。因此,學習使vHPC和mPFC之間的雙向連接強度產生了相反的變化。
2.4 學習前后,正確和錯誤執行任務時連接強度變化趨勢不同
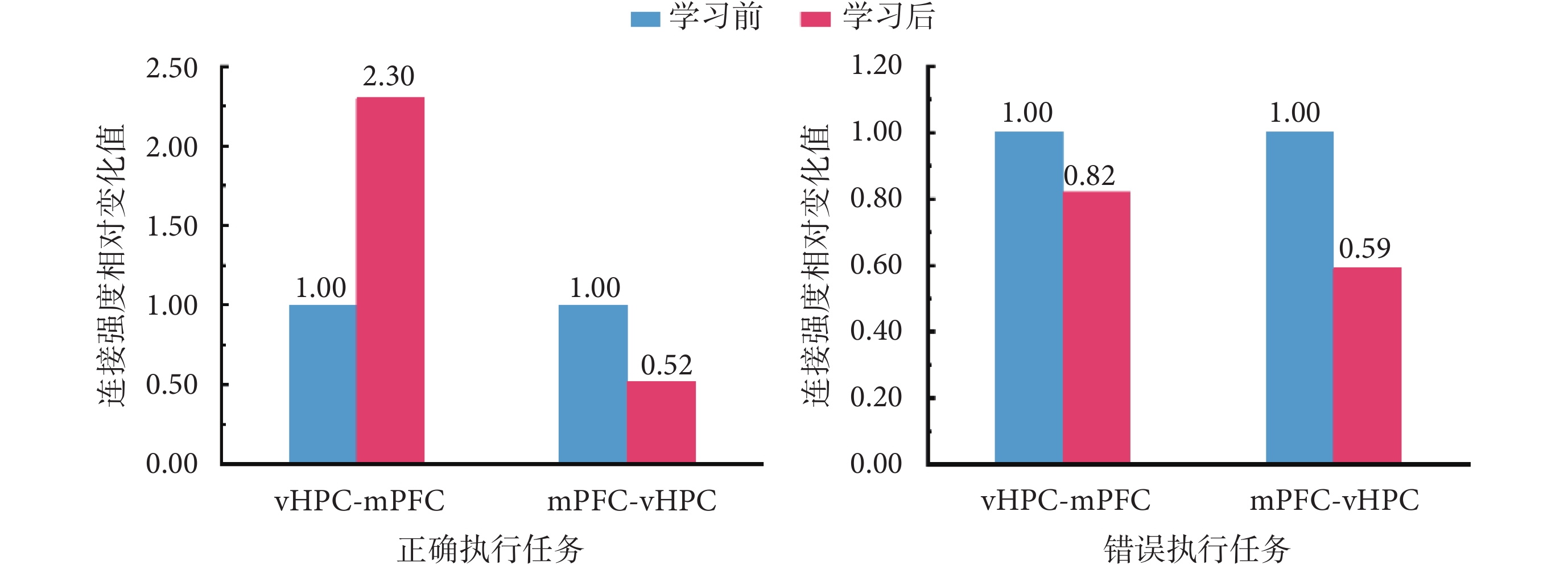
為進一步對學習前后大鼠在正確和錯誤執行任務時,兩腦區之間連接強度變化趨勢進行對比分析,以探索兩者之間是否存在差異,本文相關實驗結果如圖5所示。其中,橫坐標軸上,vHPC-mPFC,表示從vHPC到mPFC的前向連接在學習前后的對比結果;mPFC-vHPC,表示從mPFC到vHPC的后向連接在學習前后的對比結果;縱坐標軸表示,將學會前連接強度作為基準(值為1.00)時,學習前后連接強度相對變化值。
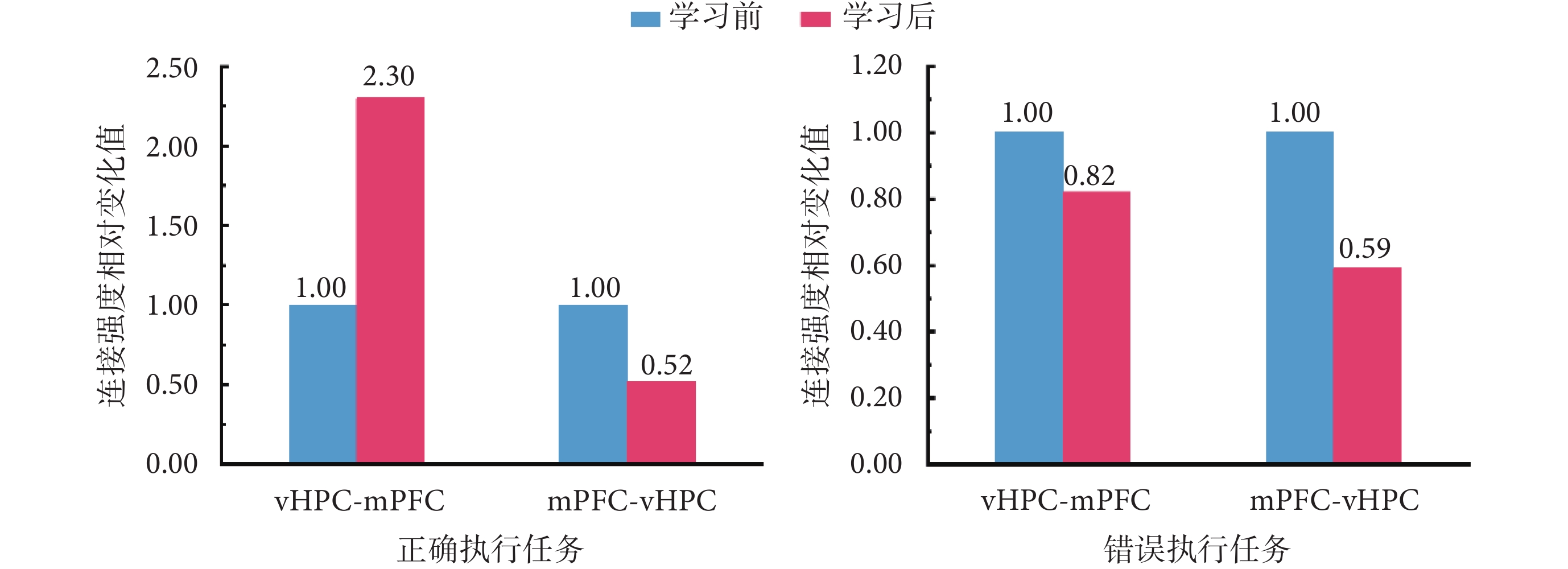 圖5
正確執行和錯誤執行 T 迷宮任務時,vHPC 和 mPFC 之間的連接強度比較
Figure5.
Comparisons of connection strength between the vHPC and mPFC for correctly and incorrectly performed T maze tasks
圖5
正確執行和錯誤執行 T 迷宮任務時,vHPC 和 mPFC 之間的連接強度比較
Figure5.
Comparisons of connection strength between the vHPC and mPFC for correctly and incorrectly performed T maze tasks
學習前后,大鼠正確執行任務時兩腦區之間連接強度對比結果顯示,相比學習前,學習后大鼠在正確執行任務時,vHPC和mPFC兩腦區之間的前向連接(vHPC-mPFC)強度增強為學習前的2.30倍,而后向連接(mPFC-vHPC)強度降低為0.52倍。此外,對大鼠學習前后,錯誤執行任務時兩腦區之間連接強度進行對比的結果則顯示,相比學習前,學習后大鼠在錯誤執行任務時,vHPC和mPFC兩腦區之間的前向連接(vHPC-mPFC)強度和后向連接(mPFC-vHPC)強度均有所下降,分別降低為學習前的0.82倍和0.59倍。對比可知,無論大鼠是否正確執行T迷宮記憶任務,與任務學習前相比,從mPFC到vHPC的后向連接強度均在學習后呈現不同程度的減弱。但是,任務學習前后,從vHPC到mPFC的前向連接強度的變化趨勢卻與大鼠是否正確執行T迷宮記憶任務密切相關。當大鼠正確執行任務時,兩腦區之間前向連接的強度增強,反之,則呈現連接強度減弱的趨勢。
3 討論與分析
vHPC和mPFC是參與學習與記憶的兩個關鍵腦區,vHPC和mPFC之間的信息傳遞在學習與記憶過程中起著重要作用[29-32]。因此,本文基于vHPC和mPFC兩個腦區的LFPs信號,應用DCM模型,探究了兩個腦區之間的連接方式以及學習對上述腦區之間連接強度的影響。結果顯示,vHPC和mPFC兩個腦區之間存在雙向連接,即從vHPC到mPFC的前向連接和從mPFC到vHPC的后向連接。該結果與此前Moran等[16]研究結果存在一致性,他們的研究對麻醉狀態下大鼠HPC和mPFC兩腦區之間的神經連接特性進行了分析,結果顯示HPC和mPFC之間也存在同樣的雙向連接。
為了進一步探究學習對mPFC和vHPC兩腦區連接強度的影響,本文中基于上述最優連接模型,進一步應用貝葉斯反演和模型的相對對數證據值對兩腦區間連接強度進行了分析。結果表明,任務學習前后,兩腦區之間的雙向連接強度均發生了改變,總體呈現從vHPC到mPFC的前向連接增強,從mPFC到vHPC的后向連接減弱的趨勢。該變化趨勢與任務學習前后,大鼠正確執行記憶任務過程中兩腦區之間連接強度的變化趨勢一致,均為前向連接增強,后向連接減弱。但是,在任務學習前后,當大鼠錯誤執行T迷宮記憶任務時,vHPC和mPFC之間的雙向連接,均呈現連接強度減弱的趨勢。上述結果表明,相比于后向連接,較高強度從vHPC到mPFC的前向連接,可能在T迷宮記憶任務的正確執行中發揮了更為重要的作用。而這種學習前后,從vHPC到mPFC前向連接的增強,則可能是由于記憶的形成和鞏固過程中突觸強度的改變和神經元興奮性的增強所引起的[33]。這與Park等[34]的研究結果存在一致性,即小鼠在學習期間執行任務正確率的提升,伴隨著vHPC神經活動的增強以及vHPC和mPFC之間連接強度的增加;此外,研究表明,由PFC向HPC的突觸投射參與了記憶鞏固和檢索等生理活動,其連接強度的變化與相關生理、病理過程密切相關。這可以作為在本研究中學習前后無論大鼠正確還是錯誤執行T迷宮記憶任務時,都存在從mPFC到vHPC的后向連接強度變化的合理解釋。而根據Rajasethupathy等[35]此前的研究,這種后向連接呈現減弱的趨勢,則可能是由于學習促使HPC中與任務相關的稀疏神經元集群的形成,使得學習后從mPFC到vHPC連接網絡的稀疏化所致。但是,目前的研究結果仍無法完全解釋,學習后,執行T迷宮記憶任務時從vHPC到mPFC的前向連接增強,而從mPFC到vHPC的后向連接減弱。此外,神經元間的連接分為興奮性連接和抑制性連接,其連接強度的改變由細胞膜上不同突觸受體介導。本文中所應用的DCM模型中,腦區間的連接強度同時受到α-氨基-3-羥基-5-甲基-4-異惡唑丙酸(α-amino-3-hydroxy-5-methyl-4-isoxazole-propionicacid,AMPA)和N-甲基-D-天冬氨酸(N-methyl-D-aspartic acid,NMDA)受體的影響。兩者在學習記憶過程中,各自對神經連接強度變化的貢獻仍有待進一步研究。
腦區間連接強度的改變由神經元上的突觸受體介導,神經連接分為興奮性連接和抑制性連接。在本文中所應用的DCM模型中,腦區間的連接為興奮性連接,其受體包括AMPA和NMDA受體,腦區內的連接包括興奮性連接和抑制性連接(其受體為γ-氨基丁酸受體)[36]。在學習記憶過程中,AMPA和NMDA受體各自對神經連接強度變化的貢獻仍有待進一步研究,以期望更加深入了解學習與記憶的神經網絡機制。
綜上,vHPC和mPFC之間因果連接特性的研究對理解學習記憶具有重要意義,本文應用DCM模型,初步探索了學習對相關腦區(vHPC和mPFC)間信息傳遞特性的影響,希望能夠為進一步從vHPC和mPFC之間神經信息網絡連接特性的角度研究學習記憶的相關機制提供支持。
重要聲明
利益沖突聲明:本文全體作者均聲明不存在利益沖突。
作者貢獻聲明:李雙燕,參與了論文的選題、實驗設計、實驗實施、論文內容審核、論文撰寫和修改;鄭衛然,參與了論文的選題、實驗設計、數據采集和分析、論文撰寫和修改;阿蘭、王龍龍、劉素紅、劉慧,參與了數據采集、分析和論文修改。
倫理聲明:本研究通過了河北工業大學生物醫學倫理委員會的批準(審查編號:HEBUTaCUC2022062)。
0 引言
學習與記憶是通過獲取、儲存和更新外部信息來實現認知活動和自我提升的關鍵[1]。學習與記憶作為大腦的高級認知活動,往往需要多個腦區協同參與才能實現[2-3]。其中,海馬(hippocampus,HPC)和前額葉皮層(prefrontal cortex,PFC)之間神經元集群活動的協同和神經信息的交互,被認為在學習與記憶功能的實現中發揮了重要作用[4-7]。例如,Morici等[8]研究發現,在對過去事件進行回憶的過程中,大鼠PFC和腹側HPC(ventral HPC,vHPC)之間的局部場電位(local field potentials,LFPs)信號同步性顯著增加。而PFC和HPC腦區之間功能連接的異常,可能是造成唐氏綜合征患者記憶功能障礙的神經機制之一[9]。此外,Avigan等[10]在大鼠十字迷宮任務的學習過程中,通過選擇性地抑制PFC或HPC中神經元的活性,發現HPC的雙側失活,嚴重損害了大鼠的空間學習和記憶功能,而雙側PFC的失活主要損害了大鼠的辨別學習能力,表明空間學習功能的實現與PFC和HPC之間的相互作用密切相關。雖然這些研究表明,HPC和PFC之間神經信息的傳遞和相互作用對于實現學習與記憶功能具有關鍵作用,但記憶相關任務學習前后,兩腦區之間的連接特性是否及如何變化,仍有待深入研究。
動態因果模型(dynamic causal modeling,DCM)是由Friston等[11]提出的一種從有效連通性角度探索腦功能相關問題的方法,目前已經在神經科學領域得到了廣泛應用[12-17]。與其它分析腦區間神經信息相互作用的方法相比,DCM模型能夠將外界環境(輸入刺激)、神經元集群的活動狀態,以及記錄所得神經電活動信號特征三者相聯立,更具有生物物理合理性[18-20]。因此,本文擬應用符合神經生理學特性的DCM算法,探究大鼠學習前后執行記憶任務過程中vHPC和內側PFC(medial PFC,mPFC)之間的因果連接特性的變化。
綜上,本文應用在體微電極記錄技術,獲取了8只大鼠在T迷宮記憶任務學習前后執行該任務過程中vHPC和mPFC腦區的LFPs信號。隨后,基于DCM模型構建了兩腦區之間可能的連接模型,并應用貝葉斯算法進行了最優模型選擇。最后,基于最優調制模型,對比分析了大鼠在T迷宮記憶任務學習前后,正確與錯誤執行該任務時vHPC和mPFC之間連接強度的變化。通過開展上述研究,本文期望能夠從腦區間因果連接特性變化的角度探索腦區間神經信息交互處理的相關機制,為今后相關領域研究奠定理論和實踐基礎。
1 材料與方法
實驗對象為8只成年雄性維斯塔爾(Wistar)大鼠,體重280~320 g,由北京華阜康生物科技股份有限公司提供。飼養期間,不限制大鼠對水和食物的獲取。任務訓練開始后,實驗動物可以獲得能夠維持其體重不低于正常體重85%的食物。本研究中的所有實驗流程與環節均嚴格遵循科研倫理規范,實驗獲河北工業大學生物醫學倫理委員會批準(審查編號:HEBUTaCUC2022062)。
1.1 T迷宮記憶任務及LFPs數據獲取
實驗開始前,需先將微電極植入大鼠mPFC和vHPC腦區。具體過程見文獻[21],簡述如下:將麻醉中的大鼠頭部固定到立體定位儀上,并使其顱骨暴露。在非目標腦區上方的顱骨固定3~4顆螺釘作為固定和接地電極。隨后,去除mPFC(前囟前2.50~4.50 mm,旁開0.20~1.00 mm)和vHPC(前囟后4.60~6.00 mm,旁開4.40~5.40 mm)上方的顱骨及硬腦膜。最后,使用微推進器P_PAD LITE(Scientifica Inc.,美國)將鎳鉻合金電極ST35(Plexon Inc.,美國)推進至目標腦區對應深度(mPFC:2.50~3.50 mm,vHPC:7.00~8.00 mm)后,使用牙科水泥進行固定。手術結束后,待大鼠恢復5~7 d,開始進行T迷宮記憶任務[22]學習并同步采集神經電生理信號。如圖1所示,T迷宮各臂寬15 cm,高20 cm,起始臂長70 cm,兩個目標臂的長度各為50 cm。T迷宮記憶任務主要包括自由選擇、延遲、交替選擇三個階段。在自由選擇階段,大鼠選擇任意目標臂均可獲得食物獎勵。隨后,經過起始點5~10 s的延遲等待后,大鼠進入交替選擇階段。在該階段,大鼠需選擇與前面自由選擇階段相異的目標臂,才可以再次得到食物獎勵,這被視為任務正確執行的標準。反之,視為錯誤執行任務。在大鼠執行任務的正確率連續2 d達到80%及以上之后,任務學習階段結束。
圖1
T迷宮記憶任務示意圖
Figure1.
Diagram of the T-maze memory task
在大鼠執行T迷宮記憶任務時應用神經電生理信號采集系統OmniPlex/128(Plexon Inc.,美國)同步記錄獲取大鼠vHPC和mPFC腦區的LFPs信號(采樣頻率1 000 Hz)。在應用DCM模型對LFPs信號進行分析之前,首先對其進行4~100 Hz的濾波以及陷波濾波,以消除50 Hz工頻干擾。
為探索學習前后大鼠在執行T迷宮記憶任務過程中,其vHPC和mPFC之間連接特性的變化,本文在后續研究中,對應任務學習前后狀態,分別選用了任務學習初始階段(開始任務訓練后第1~3天)和學習階段結束后(開始任務訓練后第8~12天),大鼠在執行任務過程中的LFPs數據進行DCM模型分析。其中,包括任務學習前:8只大鼠,共96次實驗;任務學習后:8只大鼠,共111次實驗。
1.2 DCM模型
DCM模型基于動態系統建模和貝葉斯統計框架,對神經網絡中的有效連接模式及其調節機制進行研究。該模型將大腦抽象為一個復雜的“刺激輸入—內部狀態變化—輸出響應”的動態系統,其中,外部環境的刺激作為輸入,能夠激發神經系統內部狀態的動態演變,而這些內部狀態的變化可以通過特定的輸出信號得以體現。在這一理論架構下,DCM模型巧妙利用了給定的刺激輸入與相應的輸出反饋數據,通過調整模型的參數,逆向推斷出隱藏于特定腦區背后的神經元活動變化及其相互間的有效連接模式。DCM模型中用于描述外界刺激下神經元狀態變化的狀態方程,其表達式如式(1)所示:
|
其中,?表示神經元狀態變量;F是描述外部輸入刺激u或神經元活動z對其他腦區神經元狀態變化產生生理影響的非線性函數;θ表示激活腦區生成模型中一系列待定的生物物理參數和連接參數;該方程反映了不同神經元集群之間的信息傳遞過程[23]。
此外,基于觀測方程,可以實現從目標腦區神經元集群活動狀態到測量所得信號的進一步映射[24],其表達式如式(2)所示:
|
其中,觀測信號y通過觀測函數g與隱含狀態z及觀測參數?之間建立了明確的依賴關系。
在運用DCM模型解析神經元集群間有效連接模式的過程中,確保模型的神經生物學合理性是保障分析結果準確性的前提。這要求模型參數必須富含生物物理學信息,從而增加了參數估計的復雜性與挑戰性[25]。為了高效且可靠地實現對模型參數的估計,DCM模型采用了基于貝葉斯統計學的反演方法[11]。
根據貝葉斯原理,對于特定的DCM模型m,其參數的后驗概率密度p(θ|y, m)的計算,如式(3)所示:
|
其中,p(y|θ, m)表示觀測數據y在給定參數θ與模型m條件下的似然性;p(θ|m)作為參數的先驗分布,融入了研究者對未知參數的先驗知識;而p(y|m),即模型證據值或邊際似然函數,則綜合評估了模型m對數據的整體解釋能力,其在模型選擇與評估中扮演著至關重要的角色[26]。
基于模型證據值,可以對相同樣本數據(如多腦區神經信號)所可能具有的不同連接模型的合理性進行比較[11]。模型間的比較由貝葉斯因子體現,表達式如式(4)所示:
|
其中,mi和mj分別代表兩種腦區間連接方式不同的DCM模型。貝葉斯因子BFi, j用于量化比較這兩個模型與觀測數據之間的擬合程度。具體而言,當BFi, j>1時,這指示了相較于模型mj,模型mi在解釋觀測數據方面展現出更高的證據支持,即模型mi與觀測數據的匹配性更為優越[26]。為解決由于多個貝葉斯因子相差過大而產生的難以表示的問題,可進一步對上式兩邊取對數,應用相對對數模型證據值進行表示,如式(5)所示:
|
根據相關文獻[27-28],本研究中假設同側mPFC和vHPC之間存在雙向連接。隨后,應用貝葉斯反演和模型的相對對數證據值進行最優連接模型選擇,并確定學習這一影響因素最可能對腦區間連接產生影響的情況,即最優調制模型;其中,相對對數模型證據值最大的模型,即為最優連接模型或最優調制模型。同時,基于模型后驗概率評估所選最優連接模型和調制模型的置信度,當后驗概率越接近1時,所選最優連接模型和最優調制模型的置信度越高[19]。應用上述方法,本文對比分析了大鼠學習T迷宮記憶任務前后,以及正確和錯誤執行任務時,mPFC和vHPC之間連接強度的變化,探索了學習對相關腦區神經信息傳遞的影響。本文中的DCM模型分析使用英國倫敦大學學院開發的腦成像數據序列處理軟件Statistical Parametric Mapping 12進行。
2 結果
2.1 vHPC和mPFC間最優連接模型選擇結果
如圖2所示,為vHPC和mPFC之間的連接模型及貝葉斯模型選擇結果。根據相關文獻[27],同側mPFC和vHPC之間可能存在5種不同情況的雙向連接,分別為:模型1,兩腦區之間存在雙向前向連接;模型2,兩腦區之間存在雙向后向連接;模型3,vHPC到mPFC為前向連接,mPFC到vHPC為后向連接;模型4,與前一模型相反,mPFC到vHPC為前向連接,vHPC到mPFC為后向連接;模型5,兩腦區之間同時存在雙向前向連接和雙向后向連接。本研究中,首先基于8只大鼠學習前和學習后執行T迷宮記憶任務時的LFPs數據,應用貝葉斯反演和模型的相對對數證據值進行上述模型的最優選擇。貝葉斯模型選擇結果顯示,模型3具有最大的相對對數模型證據值(157),即vHPC和mPFC之間最優的連接模型為模型3(vHPC到mPFC為前向連接,mPFC到vHPC為后向連接)。進一步對該模型的置信度進行分析的結果顯示,其后驗概率為1,模型可信。因此,接下來,進一步基于模型3,對大鼠學習前后連接受影響的可能情況進行分析。
圖2
5種源間連接類型的描述和貝葉斯模型比較結果
Figure2.
Illustration of 5 different between-source connection models and Bayesian model comparison results
2.2 最優調制模型選擇結果
基于所選擇的最優連接模型,學習這一因素對vHPC和mPFC之間連接的影響可能存在以下3種情況:一是僅vHPC到mPFC的前向連接受到影響;二是僅mPFC到vHPC的后向連接受到影響;三是兩腦區之間的雙向連接,即前向和后向連接均受到了影響。因此,所構建的連接調制模型如圖3所示,其中,虛線表示受外界因素調制的腦區間連接,實線表示未受到影響的連接。同樣,應用貝葉斯反演和模型的相對對數證據值對上述模型進行最優選擇。圖3右圖連接調制模型中,橫軸分別對應前向連接受影響、后向連接受影響、雙向連接受影響的調制模型分析結果,縱軸為相對對數模型證據值。從圖3中可以看出,雙向連接均受到影響的模型具有最大的相對對數模型證據值(5.60),是最優的調制模型。進一步對其置信度進行分析的結果表明,該模型的后驗概率為0.98,具有較高置信度(98%)。上述結果表明,對記憶任務的學習能夠同時影響vHPC和mPFC之間的雙向連接。
圖3
3種連接調制模型的描述和貝葉斯模型比較結果
Figure3.
Illustration of 3 different connection modulation models and Bayesian model comparison results
2.3 學習前后vHPC和mPFC之間雙向連接強度的變化
為研究學習前后vHPC和mPFC之間的雙向連接如何變化,本研究進一步基于任務學習前后,大鼠在執行T迷宮記憶任務過程中兩腦區的LFPs信號,應用所選最優調制模型,對任務學習前后兩腦區之間連接強度的變化進行了分析,結果如圖4所示。其中,8只大鼠每天的T迷宮記憶任務平均正確率(以均值±標準差表示)結果顯示,隨著訓練天數的增加,大鼠執行T迷宮記憶任務的平均正確率總體呈上升趨勢,最終能夠達到并維持在80%以上。如前文所述,本文中選取了每只大鼠任務學習初始階段(第1~3天)和學習階段結束后(第8~12天)的神經電活動信號進行了分析。本文基于學習前后大鼠執行T迷宮記憶任務時vHPC和mPFC的LFPs信號,應用DCM模型,對兩腦區之間連接強度變化進行了分析。圖4右圖橫坐標軸上,vHPC-mPFC,表示從vHPC到mPFC的前向連接強度在學習前后的對比結果;mPFC-vHPC,表示從mPFC到vHPC的后向連接強度在學習前后的對比結果。圖4右圖縱坐標軸,表示連接強度相對變化值。
圖4
大鼠學習 T 迷宮任務并比較 vHPC 和 mPFC 之間的連接強度
Figure4.
Changes in T maze task accuracy during rats learning and a comparison of connection strength between the vHPC and mPFC pre- and post-learning
結果顯示,相比學習前,學習后大鼠在執行任務時,vHPC和mPFC兩腦區之間的前向連接(vHPC-mPFC)增強為學習前的1.80倍,而后向連接(mPFC-vHPC)降低為0.23倍。因此,學習使vHPC和mPFC之間的雙向連接強度產生了相反的變化。
2.4 學習前后,正確和錯誤執行任務時連接強度變化趨勢不同
為進一步對學習前后大鼠在正確和錯誤執行任務時,兩腦區之間連接強度變化趨勢進行對比分析,以探索兩者之間是否存在差異,本文相關實驗結果如圖5所示。其中,橫坐標軸上,vHPC-mPFC,表示從vHPC到mPFC的前向連接在學習前后的對比結果;mPFC-vHPC,表示從mPFC到vHPC的后向連接在學習前后的對比結果;縱坐標軸表示,將學會前連接強度作為基準(值為1.00)時,學習前后連接強度相對變化值。
圖5
正確執行和錯誤執行 T 迷宮任務時,vHPC 和 mPFC 之間的連接強度比較
Figure5.
Comparisons of connection strength between the vHPC and mPFC for correctly and incorrectly performed T maze tasks
學習前后,大鼠正確執行任務時兩腦區之間連接強度對比結果顯示,相比學習前,學習后大鼠在正確執行任務時,vHPC和mPFC兩腦區之間的前向連接(vHPC-mPFC)強度增強為學習前的2.30倍,而后向連接(mPFC-vHPC)強度降低為0.52倍。此外,對大鼠學習前后,錯誤執行任務時兩腦區之間連接強度進行對比的結果則顯示,相比學習前,學習后大鼠在錯誤執行任務時,vHPC和mPFC兩腦區之間的前向連接(vHPC-mPFC)強度和后向連接(mPFC-vHPC)強度均有所下降,分別降低為學習前的0.82倍和0.59倍。對比可知,無論大鼠是否正確執行T迷宮記憶任務,與任務學習前相比,從mPFC到vHPC的后向連接強度均在學習后呈現不同程度的減弱。但是,任務學習前后,從vHPC到mPFC的前向連接強度的變化趨勢卻與大鼠是否正確執行T迷宮記憶任務密切相關。當大鼠正確執行任務時,兩腦區之間前向連接的強度增強,反之,則呈現連接強度減弱的趨勢。
3 討論與分析
vHPC和mPFC是參與學習與記憶的兩個關鍵腦區,vHPC和mPFC之間的信息傳遞在學習與記憶過程中起著重要作用[29-32]。因此,本文基于vHPC和mPFC兩個腦區的LFPs信號,應用DCM模型,探究了兩個腦區之間的連接方式以及學習對上述腦區之間連接強度的影響。結果顯示,vHPC和mPFC兩個腦區之間存在雙向連接,即從vHPC到mPFC的前向連接和從mPFC到vHPC的后向連接。該結果與此前Moran等[16]研究結果存在一致性,他們的研究對麻醉狀態下大鼠HPC和mPFC兩腦區之間的神經連接特性進行了分析,結果顯示HPC和mPFC之間也存在同樣的雙向連接。
為了進一步探究學習對mPFC和vHPC兩腦區連接強度的影響,本文中基于上述最優連接模型,進一步應用貝葉斯反演和模型的相對對數證據值對兩腦區間連接強度進行了分析。結果表明,任務學習前后,兩腦區之間的雙向連接強度均發生了改變,總體呈現從vHPC到mPFC的前向連接增強,從mPFC到vHPC的后向連接減弱的趨勢。該變化趨勢與任務學習前后,大鼠正確執行記憶任務過程中兩腦區之間連接強度的變化趨勢一致,均為前向連接增強,后向連接減弱。但是,在任務學習前后,當大鼠錯誤執行T迷宮記憶任務時,vHPC和mPFC之間的雙向連接,均呈現連接強度減弱的趨勢。上述結果表明,相比于后向連接,較高強度從vHPC到mPFC的前向連接,可能在T迷宮記憶任務的正確執行中發揮了更為重要的作用。而這種學習前后,從vHPC到mPFC前向連接的增強,則可能是由于記憶的形成和鞏固過程中突觸強度的改變和神經元興奮性的增強所引起的[33]。這與Park等[34]的研究結果存在一致性,即小鼠在學習期間執行任務正確率的提升,伴隨著vHPC神經活動的增強以及vHPC和mPFC之間連接強度的增加;此外,研究表明,由PFC向HPC的突觸投射參與了記憶鞏固和檢索等生理活動,其連接強度的變化與相關生理、病理過程密切相關。這可以作為在本研究中學習前后無論大鼠正確還是錯誤執行T迷宮記憶任務時,都存在從mPFC到vHPC的后向連接強度變化的合理解釋。而根據Rajasethupathy等[35]此前的研究,這種后向連接呈現減弱的趨勢,則可能是由于學習促使HPC中與任務相關的稀疏神經元集群的形成,使得學習后從mPFC到vHPC連接網絡的稀疏化所致。但是,目前的研究結果仍無法完全解釋,學習后,執行T迷宮記憶任務時從vHPC到mPFC的前向連接增強,而從mPFC到vHPC的后向連接減弱。此外,神經元間的連接分為興奮性連接和抑制性連接,其連接強度的改變由細胞膜上不同突觸受體介導。本文中所應用的DCM模型中,腦區間的連接強度同時受到α-氨基-3-羥基-5-甲基-4-異惡唑丙酸(α-amino-3-hydroxy-5-methyl-4-isoxazole-propionicacid,AMPA)和N-甲基-D-天冬氨酸(N-methyl-D-aspartic acid,NMDA)受體的影響。兩者在學習記憶過程中,各自對神經連接強度變化的貢獻仍有待進一步研究。
腦區間連接強度的改變由神經元上的突觸受體介導,神經連接分為興奮性連接和抑制性連接。在本文中所應用的DCM模型中,腦區間的連接為興奮性連接,其受體包括AMPA和NMDA受體,腦區內的連接包括興奮性連接和抑制性連接(其受體為γ-氨基丁酸受體)[36]。在學習記憶過程中,AMPA和NMDA受體各自對神經連接強度變化的貢獻仍有待進一步研究,以期望更加深入了解學習與記憶的神經網絡機制。
綜上,vHPC和mPFC之間因果連接特性的研究對理解學習記憶具有重要意義,本文應用DCM模型,初步探索了學習對相關腦區(vHPC和mPFC)間信息傳遞特性的影響,希望能夠為進一步從vHPC和mPFC之間神經信息網絡連接特性的角度研究學習記憶的相關機制提供支持。
重要聲明
利益沖突聲明:本文全體作者均聲明不存在利益沖突。
作者貢獻聲明:李雙燕,參與了論文的選題、實驗設計、實驗實施、論文內容審核、論文撰寫和修改;鄭衛然,參與了論文的選題、實驗設計、數據采集和分析、論文撰寫和修改;阿蘭、王龍龍、劉素紅、劉慧,參與了數據采集、分析和論文修改。
倫理聲明:本研究通過了河北工業大學生物醫學倫理委員會的批準(審查編號:HEBUTaCUC2022062)。