引用本文: 符培亮, 叢銳軍, 陳松, 張雷, 丁喆如, 周琦, 李林濤, 許震宇, 吳宇黎, 吳海山. 滑膜間充質干細胞成纖維軟骨分化條件初步探索. 中國修復重建外科雜志, 2015, 29(1): 81-91. doi: 10.7507/1002-1892.20150018 復制
版權信息: ?四川大學華西醫院華西期刊社《中國修復重建外科雜志》版權所有,未經授權不得轉載、改編
半月板損傷后再生能力有限,特別是白區,長期失去半月板保護會導致膝關節疼痛、功能障礙甚至殘疾。近年組織工程學的發展為半月板修復提供了新方法。滑膜間充質干細胞(synovial derived MSCs,SMSCs)取材簡便、對患者創傷小、細胞量大、成纖維軟骨分化能力強,有望成為理想的種子細胞來源。體外誘導SMSCs成纖維軟骨分化的條件尚未明確[1-5]。本實驗探索了誘導SMSCs成纖維軟骨分化的最佳條件,觀察不同誘導條件對SMSCs成纖維軟骨分化的影響以及不同誘導條件對分化后細胞表型的影響,以期為SMSCs體外誘導構建組織工程半月板奠定理論基礎。
1 材料與方法
1.1 實驗動物及主要試劑、儀器
15月齡雄性新西蘭白兔5只,體質量(2.20 ±0.65)kg,由第二軍醫大學動物中心提供。
0.05 %胰蛋白酶-0.02%EDTA(- 20℃保存;GIBCO 公司,美國);10 ng/mL TGF-β3、500 ng/ mL BMP-2、100 nmol/L 地塞米松(dexamethasone,DEX)、40 μg/mL脯氨酸、50 μg/mL檸檬酸(ascorbic acid,ASA)、100 μg/mL丙酮酸、6.25 μg/ mL胰島素、6.25 μg/mL轉鐵蛋白、6.25 μg/mL亞硒酸、1.25 mg/ mL牛血清白蛋白(bovine serum albumin,BSA)、5.35 mg/ mL 亞麻酸、10 ng/mL重組人TGF-β1、50 ng/ mL bFGF、1.2%藻酸鹽溶液、50 ng/mL BMP-7、10 ng/mL IGF、10 ng/mL TGF-β2、50 ng/mL BMP-4、Taq DNA聚合酶、10×PCR緩沖液、dNTP +熒光酶(Sigma公司,美國);PBS緩沖液、0.01 mol/ L檸檬酸鹽緩沖液、0.5 mol/L EDTA緩沖液、1 mol/L TBS緩沖液、4%多聚甲醛、10%山羊血清、3%甲醇-H 2O2溶液(上海試劑廠);鼠抗兔Ⅰ型膠原(collagen typeⅠ,ColⅠ)、Ⅱ型膠原蛋白一抗、鼠抗兔CD105一抗(BioLegend公司,美國);FITC標記的馬抗鼠二抗、TRITC標記的馬抗鼠二抗、鼠抗兔CD44一抗(BD Pharmingen公司,美國)。
5060/BB16自動溫控CO2培養箱(Heraeus公司,德國);壓力控制外設(第二軍醫大學組胚實驗室改裝);超凈臺(蘇州凈化設備有限公司);HIMAC CR21高速冷凍離心機(Hitachi公司,日本);低速離心機(Thermo公司,美國);倒置相差顯微鏡、HFX-DX顯微攝影系統(Nikon公司,日本);超低溫冰箱(Forma Scientific公司,美國);Labconco Water PROTMPS超純水制備系統(Labconco公司,美國);微量加樣器(Gilson公司,法國);一次性培養瓶、培養皿及培養板、10 mL/5 mL一次性吸管、凍存管、離心管、細胞培養皿(Geriner&Corning公司,美國);程序降溫盒(Nalgen公司,美國);FACSCalibur 流式細胞儀、CellQuest 軟件系統(Becton Dickinson公司,美國)。
1.2 SMSCs分離培養及鑒定
1.2.1 SMSCs分離培養
無菌條件下取新西蘭白兔膝關節1 cm×1 cm滑膜組織,無菌PBS溶液反復沖洗3次,加入1%(V/V)抗生素(10 000 U/mL青霉素、10 000 μg/mL鏈霉素、25 μg/mL兩性霉素B)。于DMEM中加入3 mg/mL膠原蛋白酶Ⅱ消化組織塊。37℃、5%CO2孵育過夜,40 μm尼龍細胞濾器移除不溶物。4℃以150 × g離心7 min,DMEM沖洗2遍后重懸收獲細胞;37℃、5%CO2培養箱中培養72 h,移除未貼壁細胞,加入含10%(V/V)小牛血清的α-MEM培養基培養細胞,每周換液2次,待細胞長滿培養皿后,0.05%胰蛋白酶消化并傳代,倒置相差顯微鏡下觀察細胞形態。
1.2.2 細胞擴增分析
SMSCs培養期間每隔3 d檢測細胞擴增情況。采用Proteinase K試劑盒提取細胞DNA,PicoGreen Assay測量細胞總DNA量,通過DNA量反映細胞擴增情況,繪制細胞擴增曲線。
1.2.3 SMSCs鑒定
① 流式細胞術:根據文獻[2]方法采用流式細胞儀鑒定原代貼壁細胞表面標志物CD44、CD90、CD105、CD14、CD34和CD45表達。
② 成軟骨分化:取20 μL第4代SMSCs,調整細胞濃度為2×107個/mL,接種于24孔板,貼壁3 h后加入成軟骨誘導培養液(含10 ng/mL TGF-β1、50 μg/ mL抗壞血酸、6.25 μg/mL胰島素、6.25 μg/ mL轉鐵蛋白、6.25 ng/mL亞硒酸、1.25 mg/mL BSA、5.35 mg/mL亞麻酸、100 nmol/L DEX的DMEM培養基),每2天更換培養基,維持TGF-β1濃度為10 ng/mL。培養14 d后PBS沖洗,- 20℃甲醇固定30 min,甲苯胺藍染色過夜,蒸餾水反復沖洗,以200 μL 6 mol/L鹽酸胍室溫下萃取,分光光度法測定波長630 nm處吸光度(A)值。
③ 成骨分化:如前步驟于24孔板內加入成骨誘導培養基(含100 nmol/L DEX、0.2 mmol/L ASA、10 mmol/L β-甘油磷酸鈉的DMEM培養基),培養21 d后加入0.2 nmol/L CaCl2,PBS沖洗后,4%多聚甲醛固定,2%茜素紅染色(pH4.2)3 min,蒸餾水沖洗,倒置相差顯微鏡下觀察。
④ 成脂分化:根據文獻[4]方法進行成脂分化鑒定。如前步驟于24孔板內加入成脂誘導培養基(含1 μmol/L DEX、0.5 mmol/L 3-異丁基-1-甲基黃嘌呤、100 μmol/L吲哚美辛、10 μg/mL胰島素的DMEM培養基)。誘導21 d后4%多聚甲醛固定1 h,0.5%油紅O染色2 h,倒置相差顯微鏡下觀察。
1.3 缺失實驗設計
首先使用缺失實驗方法,探索SMSCs成纖維軟骨分化的必要條件,再根據缺失實驗結果設計正交實驗。缺失實驗實驗條件包括:10 ng/ mL TGF-β3、500 ng/ mL BMP-2、100 nmol/L DEX、40 μg/mL 脯氨酸、50 μg/ mL ASA、100 μg/mL丙酮酸、6.25 μg/mL胰島素、6.25 μg/ mL轉鐵蛋白、6.25 μg/mL亞硒酸、1.25 mg/ mL BSA、5.35 mg/mL亞麻酸、10 ng/mL重組人TGF-β1、50 ng/mL bFGF、1.2%藻酸鹽溶液、循環間斷靜水壓(intermittent hydraulic pressure,IHP;0.5 kPa、1 Hz)、低氧環境(2%O2)、50 ng/mL BMP-7、10 ng/ mL IGF、10 ng/mL TGF-β2、50 ng/mL BMP-4、37℃、三維環境、5%CO2。上述條件中,三維環境采用小腸黏膜下層(small intestinal submucosa,SIS)支架,三維條件缺失時采用凝膠支架,實現單層培養。為明確溫度對SMSCs成纖維軟骨分化的影響,設立獨立對照實驗:采用隨機數字表法定義46.4、4.5、20.0℃作為溫度對照,并采用30~40℃連續0.5℃階梯變溫,探索溫度對SMSCs成纖維軟骨分化能力的調控作用。
取第4代SMSCs以2.5 ×105個/mL按上述實驗條件誘導,每組缺失1個條件(1~23組,見表 1),以加入所有誘導條件為陽性對照組(24組)。倒置相差顯微鏡下觀察各組細胞形態;培養14 d甲苯胺藍染色,倒置相差顯微鏡下觀察支架復合體形態及染色情況;并行半定量RT-PCR和免疫組織化學染色測定各組細胞是否表達SOX9、聚集蛋白聚糖(Aggrecan,AGN)、ColⅠ、ColⅡ以及ColⅨ。① 半定量RT-PCR:使用RNeasy Mini Kit試劑盒提取總RNA,使用SuperScriptⅢFirst-Strand Synthesis System Kit試劑盒逆轉錄得到cDNA,采用半定量RT-PCT對目的基因進行擴增,引物序列見表 2;在擴增進入平臺期前,檢測各孔A值,對基因濃度進行半定量分析,數據僅用于輔助分析成軟骨分化趨勢。② 免疫組織化學染色:采用免疫熒光法檢測高表達ColⅠ的細胞占軟骨細胞比例。封閉抗原后,采用鼠抗兔ColⅠ、ColⅡ一抗,連接了熒光素酶的馬抗鼠二抗,使用FITC和TRITC顯色。手工板計數,隨機取10個不同視野下10個大格內全部軟骨細胞作為總體,由于彈性軟骨細胞數量稀少,所以高表達ColⅠ的纖維軟骨細胞與高表達ColⅡ的透明軟骨細胞之和應等于軟骨細胞總體;高表達ColⅠ的纖維軟骨細胞計數與該總體比值視為纖維軟骨轉化率(N),以10個視野下各轉化單元N值的復合標準差作為實驗的總體標準差。實驗重復3次。
引入rate作為缺失實驗檢驗指標,rate定義為CD151+/CD44+細胞數與N值的乘積;再進一步結合細胞形態、甲苯胺藍染色、半定量RT-PCR檢測SOX9、AGN、ColⅠ、ColⅡ、ColⅨ基因表達來驗證rate值的信度。SMSCs成纖維軟骨分化后流式細胞計數和染色得到的N值必須與半定量RT-PCR檢測的基因表達趨勢一致,rate值才能用于統計分析,差異表達的結果不會出現在結果部分。為提高實驗信度,實驗平行3次,結果中報道的轉化值為最有代表性的實驗結果,而不是3次實驗的均值,存在交互作用的因子進一步行相關性分析,成直線正相關的兩個因子僅保留N值相對較高的因子。
1.4 正交實驗設計
根據缺失實驗篩選出調控條件:TGF-β1、BMP-2、DEX、脯氨酸、ASA、丙酮酸、胰島素+轉鐵蛋白+亞硒酸預混液(insulin + transferring + seleniousacid,ITS)、BSA、bFGF、IHP、BMP-7、IGF對SMSCs成纖維軟骨調節作用顯著(表 3),納入2水平正交實驗,各因子的水平參考文獻[1-5]獲得;溫度、CO2濃度和低氧環境存在交互作用,這些條件改變或缺失后SMSCs不能成纖維軟骨分化。其中,ITS預混溶液中各因子對SMSCs成纖維軟骨分化的影響存在直線正相關,此處將胰島素、轉鐵蛋白和亞硒酸作為一個因素設計為ITS。根據我們的實驗條件可進行單組<64孔的培養,選擇L60(212)正交設計能夠滿足實驗且最大限度保證結果準確性。使用SPSS18.0統計軟件設計正交實驗表頭(表 4),定義兩個水平值為水平1、水平2;根據表 1中的不同實驗條件對SMSCs行成纖維軟骨誘導。采用流式細胞儀計數,檢測CD151+/CD44+細胞并記錄N值;采用免疫組織化學染色,結合細胞形態、甲苯胺藍染色、半定量RT-PCR檢測SOX9、AGN、ColⅠ、ColⅡ、ColⅨ基因表達,進一步驗證結果。檢驗指標rate是CD151+/CD44+細胞與N值的乘積,計算方法同缺失實驗。
1.5 統計學方法
為防止不同個體來源SMSCs成纖維軟骨分化差異影響,采用來源于同一動物的SMSCs重復3 次實驗,取均值和標準差,并計算95%可信區間。來源于其余4只實驗動物的SMSCs作為對照,只有5組同時得到有意義的陽性結果,且結果趨勢一致,才能作為實驗結果,結果部分報道的N值是最典型的1只實驗動物的數據。缺失實驗采用方差分析,比較各組與標準條件組間的關系,排除非必要的調控因子;正交實驗結果采用直觀觀察和主體間方差分析方法,考慮部分因子間的1階交互作用,組間比較采用LSD和q檢驗,采用Ⅲ型平方和校正模型;檢驗水準α=0.05。
2 結果
2.1 SMSCs形態學觀察及鑒定
2.1.1 SMSCs形態學觀察及細胞擴增結果
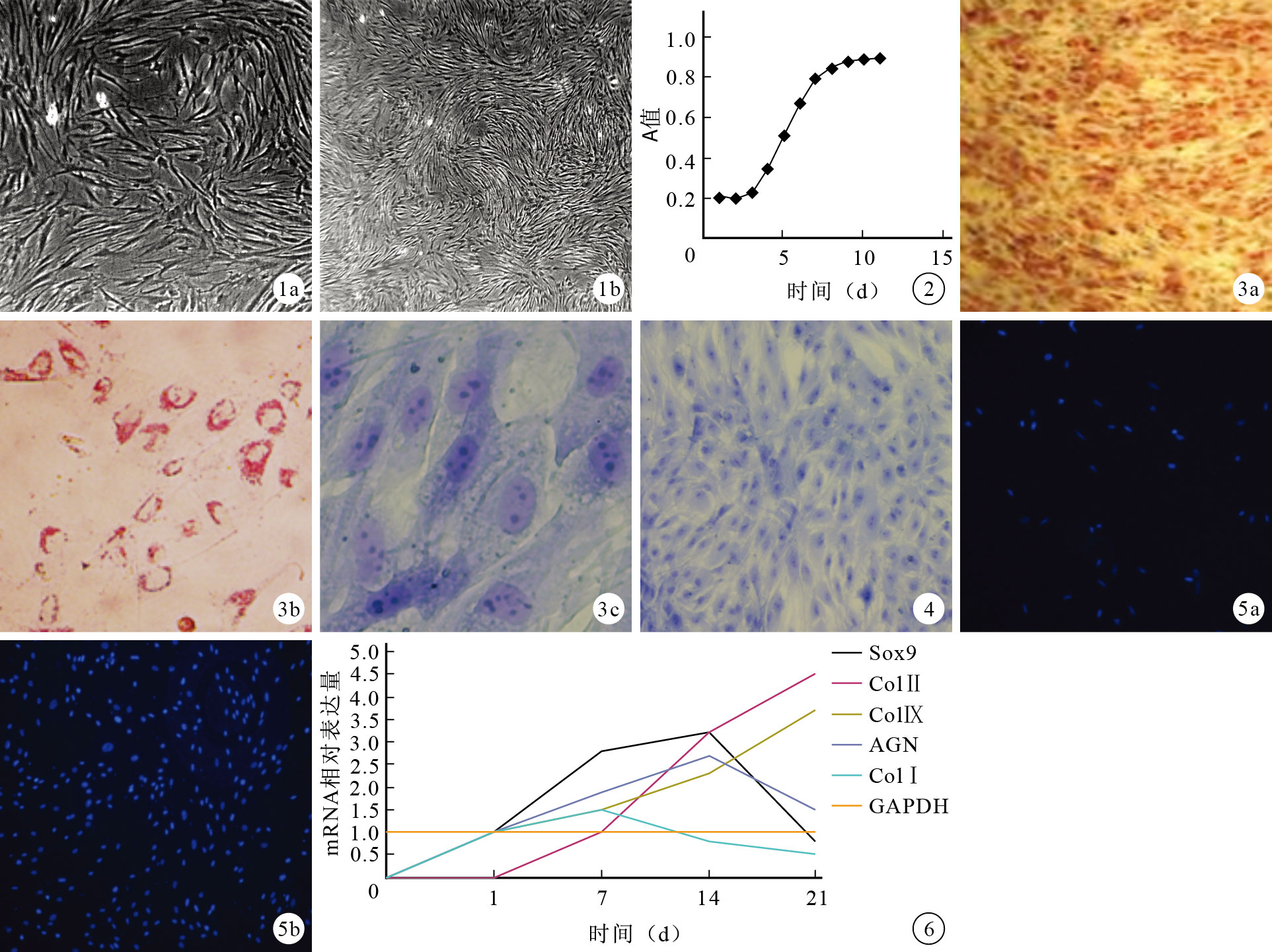
原代細胞呈多邊形或星形;第1代細胞形態逐漸變成圓形和梭形;隨著傳代代次增加,細胞形態逐漸均一,呈“向日葵”樣分布,第3代細胞呈梭形。見圖 1。細胞倍增時間為28 h,細胞擴增曲線見圖 2。
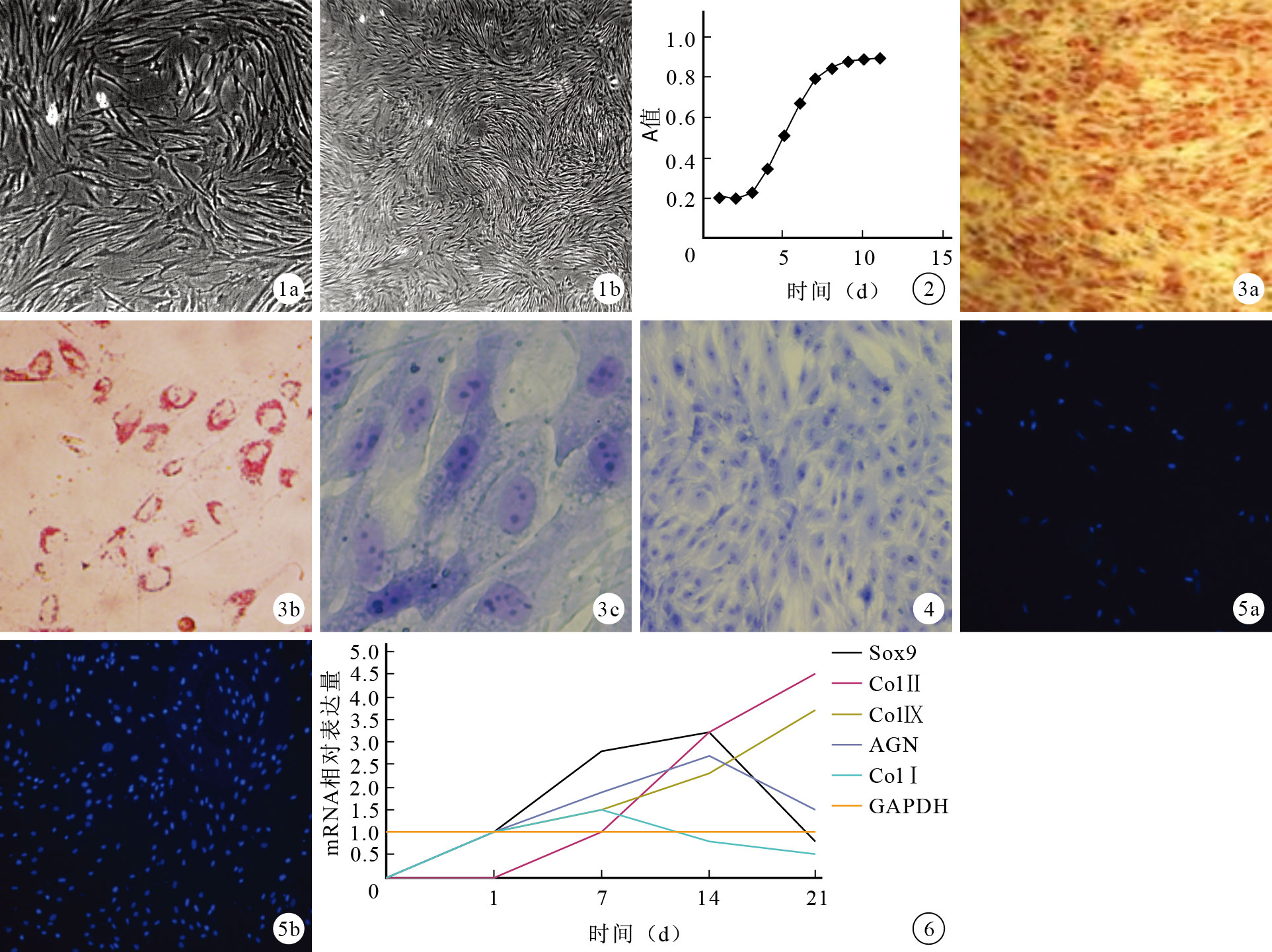 圖1
SMSCs形態學觀察(倒置相差顯微鏡×40) ? 原代細胞 ? 第3代細胞 ? ?
圖1
SMSCs形態學觀察(倒置相差顯微鏡×40) ? 原代細胞 ? 第3代細胞 ? ?2.1.2 SMSCs鑒定
流式細胞儀檢測示SMSCs表面標志CD44、CD90、CD105呈陽性表達,CD14、CD34、CD45呈陰性表達。多向分化鑒定示,成骨誘導21 d茜素紅染色、成軟骨誘導14 d甲苯胺藍染色及成脂誘導21 d油紅O染色均呈陽性(圖 3)。
2.2 SMSCs體外成軟骨分化觀察
按照正交實驗培養條件誘導3 d后,細胞聚集成小團狀,其趨化因子不清楚,細胞沿支架間隙爬行生長;支架體積在培養5 d倍增;14 d細胞-支架復合物甲苯胺藍染色呈陽性(圖 4)。軟骨細胞免疫組織化學染色示ColⅠ、ColⅡ呈陽性染色(圖 5)。半定量RT-PCR檢測示,誘導后細胞表達SOX9、AGN、ColⅠ、ColⅡ、ColⅨ基因(圖 6),各組基因表達不同,誤差線表示50%可信區間。
2.3 缺失實驗結果
缺失實驗結果顯示,TGF-β1、BMP-2、DEX、脯氨酸、抗壞血酸、丙酮酸、ITS、BSA、bFGF、IHP、BPM-7、IGF對SMSCs成纖維軟骨分化調控作用顯著(表 5),且各因子間不存在交互作用,作為獨立因子參與正交實驗。溫度降低、升高,高氧環境以及使用凝膠限制細胞接觸的狀態下,細胞誘導分化明顯延遲。來源于同一細胞株的SMSCs,限制細胞接觸后成纖維軟骨分化能力不同;溫度對SMSCs成纖維軟骨分化的影響并非成線性,在35.5~38.5℃范圍內,成纖維軟骨分化能力無顯著差異,當溫度高于或低于此范圍時,SMSCs成纖維軟骨分化能力下降。因此,溫度、氧環境以及細胞間接觸是SMSCs成纖維軟骨分化必不可少的條件,但條件本身在閾值范圍內改變時影響并不顯著,超過閾值后SMSCs幾乎不能成纖維軟骨分化,因此不作為正交實驗因子。亞麻酸缺失對成纖維軟骨分化影響不顯著,且與其他因子不存在交互作用,為非必要調控因素;方差分析顯示TGF-β1、TGF-β3與 TGF-β2存在交互作用,成線性正相關,且不與其他因子交互,TGF-β1、TGF-β2與TGF-β3同時缺失,SMSCs不能成纖維軟骨分化,缺失TGF-β1時SMSCs成纖維軟骨分化能力較強,但纖維軟骨細胞分化率僅為1.18%,而缺失TGF-β3、TGF-β2時為61.89%和75.13%,因此保留TGF-β1作為調控因子。藻酸鹽缺失狀態下,SMSCs成纖維軟骨分化率為62.64%,與陽性對照組間比較差異無統計學意義(P>0.05),且與各指標間無交互作用,為非必要調控因素。
2.4 正交實驗結果
正交實驗結果見表 6,直觀分析顯示,按轉化率降序擴展排序后,取轉化率中位數為分界線區組觀察,第1因子TGF-β1對轉化率的作用最明顯,TGF-β1高濃度組轉化率均>30%、低濃度組轉化率<30%;其余分界線兩側的水平值加和均為45,直觀條件下無法判斷其轉化率差異。取轉化率1/4位數區組觀察,見第11因子BMP-7第1區組轉化率最高,水平值加和顯示第1區組為30,第2、3、4區組(轉化率依次降低)水平值加和分別為15、23、22,說明BMP-7采用水平2有利于得到更高的纖維軟骨轉化率;其余第1區組水平值加和高于22.5的因子有第3、5、7、9、12因子,第4區組的水平值加和均低于22.5,說明采用本正交實驗表頭設計的正交實驗,各因子與轉化率之間有較好的相關性。
正交實驗方差分析采用直觀觀察和主體間方差分析的方法,結果見表 6。校正模型P=0.000,表明該模型能夠滿足實驗設計;截距P=0.000,表明各因子對因變量影響差異不完全相同。TGF-β1、ASA、bFGF、IGF、BMP-7等因子對因變量調控作用較其他因子顯著,差異有統計學意義;即改變以上因子濃度,SMSCs成纖維軟骨分化能力顯著降低,上述因子是SMSCs成纖維軟骨分化最重要的調節因素,與直觀觀察結果相似。
3 討論
本研究結果表明,SMSCs成纖維軟骨分化過程中,TGF-β1、ASA、bFGF、IGF 4個因子劑量的改變顯著影響SMSCs成纖維軟骨分化作用,通過合理調整上述因子濃度,可顯著提高成纖維軟骨細胞分化轉化率;但由于檢索資源限制,我們的實驗設計不可能覆蓋所有敏感因子,這4個因子并不是SMSCs成纖維軟骨分化唯一調控因素。TGF-β是公認的SMSCs向軟骨細胞分化最關鍵的啟動因素[1-2],但對不同亞型的TGF-β促MSCs成纖維軟骨分化作用存在爭議[3-5]。Kim等[5]指出瞬間暴露在高劑量TGF-β3環境中,MSCs功能和表型被誘導變異,并能在體內增強軟骨修復能力。Vanneaux等[3]認為TGF-β受體Ⅱ在TGF-β1介導的MSCs向軟骨細胞分化過程中至關重要,TGF-β1是介導SMSCs成纖維軟骨分化的最重要因子。我們的實驗表明,不同亞型的TGF-β誘導MSCs成纖維軟骨分化能力不同,TGF-β1調節作用最強。胰島素是MSCs成纖維軟骨分化中必要的調節因子[6-7],但本實驗結果表明胰島素對SMSCs成纖維軟骨分化的調節作用較弱,指數級的劑量差異仍不能引起纖維軟骨轉化率的差異,且與TGF和亞硒酸之間存在線性交互作用,不能作為獨立的調節因子。
BMP亦是重要的纖維軟骨生成調節因子[8],許多文獻定義BMP為促進劑[9],但是對各亞型BMP作用存在爭議[10-11]。我們的實驗表明BMP-2促SMSCs成纖維軟骨分化能力最強,已經明確BMP-2調節通路是:BMP-2→TGF-β受體Ⅱ→Smad 1、5、8通路,可能由于受體的飽和作用,BMP-2調控存在天花板效應[12],高于調控濃度上限的BMP-2不會增加SMSCs成纖維軟骨分化能力,我們的實驗結果表明500 ng/ mL BMP-2與100 ng/mL BMP-2間差異不顯著,但是與缺失組對照,其促軟骨生成能力顯著提高。此外,BMP-2促MSCs成纖維軟骨分化作用機制尚存在爭議。壓力作為獨立調控因素能夠通過調節內源性BMP-2表達來調控MSCs成纖維軟骨分化過程;同時,壓力因素與BMP-2存在交互作用,Valhmu等[13]實驗證實壓力具有促進MSCs成纖維軟骨分化和促進軟骨細胞分泌AGN的作用。Wong等[14]研究也證實了壓力能夠促進軟骨細胞基質分泌。Feng等[15-16]認為鼠類BMP-2、4的啟動子區域存在與壓力應答共存的區域,這可能是壓力調控的基因基礎。Rui等[17]、Lui等[18]、Ando等[19]均認為細胞外基質的破壞和重構與該過程密切相關。Chikazu等[20]提出了BMP-2→COX2→前列腺素E2(prostaglandin E2,PGE2)(Cbfa)→骨/軟骨形成的調控模型,認為PGF2存在對BMP-2的抑制作用,Cbfa基因的高表達促進成骨作用,抑制成軟骨作用,可能與BMP-2濃度的天花板效應相關。Siddhivarn等[21]提出的Δ12-PGF J2/過氧化物酶體增殖物激活受體-γ2/BMP-2通路可能是壓力與BMP-2相關的調節通路。
ASA在MSCs成骨分化中的作用及調控通路已被廣泛研究[22-23],在成纖維軟骨分化過程中,ASA是MSCs重要細胞因子,但其在成纖維軟骨分化過程中的調控作用研究較少[24]。關于ASA和SMSCs間相互作用的報道,在筆者委托的查新報告中未見收獲。我們的實驗表明,ASA在SMSCs成纖維軟骨分化過程中可作為單獨的調控因子,且與其他因子不存在交互作用。bFGF促進MSCs成軟骨分化的作用是明確的[25],其調節通路主要通過抑制α-平滑肌動蛋白的表達,維持成軟骨必要的三維形態,從而促進MSCs成軟骨分化過程。Kim等[26]研究證實,10 ng/ mL bFGF在成纖維軟骨前的預處理能顯著促進SMSCs成纖維軟骨分化作用;值得注意的是,同組研究者在2009年卻發布了截然相反的報道,Lee等[27]認為加入20 ng/mL bFGF和血清后,成纖維軟骨分化過程受到顯著抑制;這一因子的調節通路至今尚未得到深入研究。我們的實驗結果表明,50 ng/ mL bFGF較空白對照組顯著增強了SMSCs成纖維軟骨分化能力,成透明軟骨能力顯著下降,但在加入高濃度(200 ng/ mL)bFGF后,SMSCs成纖維軟骨和透明軟骨能力均顯著下降;針對bFGF可能調節通路的下游因子實驗,我們在5組動物實驗中得到矛盾的結果,故未在本文報道,目前我們仍不能確定該因子的作用機制,將在后續研究中進一步探討。
其他作用因子對MSCs成纖維軟骨分化的作用存在爭議,如DEX作為獨立調節因子作用不穩定,Nogami等[9]報道DEX能夠增強人MSCs成纖維軟骨分化過程,但Liu等[28]報道認為無論是否添加DEX,TGF-β2均能很好誘導成軟骨,我們的研究同樣表明,DEX作為獨立因子,對SMSCs成纖維軟骨調控能力不顯著,少量DEX(<100 μg/ mL)能促進SMSCs成纖維軟骨分化,但大劑量DEX(>1 000 μg/ mL)對SMSCs成脂分化表現出更強的促進作用。此外,我們觀察到擴增階段培養基中糖和血清的濃度對成軟骨分化存在調控作用,目前未見相關報道,這一作用有待深入研究。
綜上述,本文采用正交實驗方法,探索了SMSCs成纖維軟骨分化條件,找到了效率相對較高的SMSCs成纖維軟骨分化實驗條件,但細胞生長是一動態變化過程,許多條件的時間相關性還需深入探索。
半月板損傷后再生能力有限,特別是白區,長期失去半月板保護會導致膝關節疼痛、功能障礙甚至殘疾。近年組織工程學的發展為半月板修復提供了新方法。滑膜間充質干細胞(synovial derived MSCs,SMSCs)取材簡便、對患者創傷小、細胞量大、成纖維軟骨分化能力強,有望成為理想的種子細胞來源。體外誘導SMSCs成纖維軟骨分化的條件尚未明確[1-5]。本實驗探索了誘導SMSCs成纖維軟骨分化的最佳條件,觀察不同誘導條件對SMSCs成纖維軟骨分化的影響以及不同誘導條件對分化后細胞表型的影響,以期為SMSCs體外誘導構建組織工程半月板奠定理論基礎。
1 材料與方法
1.1 實驗動物及主要試劑、儀器
15月齡雄性新西蘭白兔5只,體質量(2.20 ±0.65)kg,由第二軍醫大學動物中心提供。
0.05 %胰蛋白酶-0.02%EDTA(- 20℃保存;GIBCO 公司,美國);10 ng/mL TGF-β3、500 ng/ mL BMP-2、100 nmol/L 地塞米松(dexamethasone,DEX)、40 μg/mL脯氨酸、50 μg/mL檸檬酸(ascorbic acid,ASA)、100 μg/mL丙酮酸、6.25 μg/ mL胰島素、6.25 μg/mL轉鐵蛋白、6.25 μg/mL亞硒酸、1.25 mg/ mL牛血清白蛋白(bovine serum albumin,BSA)、5.35 mg/ mL 亞麻酸、10 ng/mL重組人TGF-β1、50 ng/ mL bFGF、1.2%藻酸鹽溶液、50 ng/mL BMP-7、10 ng/mL IGF、10 ng/mL TGF-β2、50 ng/mL BMP-4、Taq DNA聚合酶、10×PCR緩沖液、dNTP +熒光酶(Sigma公司,美國);PBS緩沖液、0.01 mol/ L檸檬酸鹽緩沖液、0.5 mol/L EDTA緩沖液、1 mol/L TBS緩沖液、4%多聚甲醛、10%山羊血清、3%甲醇-H 2O2溶液(上海試劑廠);鼠抗兔Ⅰ型膠原(collagen typeⅠ,ColⅠ)、Ⅱ型膠原蛋白一抗、鼠抗兔CD105一抗(BioLegend公司,美國);FITC標記的馬抗鼠二抗、TRITC標記的馬抗鼠二抗、鼠抗兔CD44一抗(BD Pharmingen公司,美國)。
5060/BB16自動溫控CO2培養箱(Heraeus公司,德國);壓力控制外設(第二軍醫大學組胚實驗室改裝);超凈臺(蘇州凈化設備有限公司);HIMAC CR21高速冷凍離心機(Hitachi公司,日本);低速離心機(Thermo公司,美國);倒置相差顯微鏡、HFX-DX顯微攝影系統(Nikon公司,日本);超低溫冰箱(Forma Scientific公司,美國);Labconco Water PROTMPS超純水制備系統(Labconco公司,美國);微量加樣器(Gilson公司,法國);一次性培養瓶、培養皿及培養板、10 mL/5 mL一次性吸管、凍存管、離心管、細胞培養皿(Geriner&Corning公司,美國);程序降溫盒(Nalgen公司,美國);FACSCalibur 流式細胞儀、CellQuest 軟件系統(Becton Dickinson公司,美國)。
1.2 SMSCs分離培養及鑒定
1.2.1 SMSCs分離培養
無菌條件下取新西蘭白兔膝關節1 cm×1 cm滑膜組織,無菌PBS溶液反復沖洗3次,加入1%(V/V)抗生素(10 000 U/mL青霉素、10 000 μg/mL鏈霉素、25 μg/mL兩性霉素B)。于DMEM中加入3 mg/mL膠原蛋白酶Ⅱ消化組織塊。37℃、5%CO2孵育過夜,40 μm尼龍細胞濾器移除不溶物。4℃以150 × g離心7 min,DMEM沖洗2遍后重懸收獲細胞;37℃、5%CO2培養箱中培養72 h,移除未貼壁細胞,加入含10%(V/V)小牛血清的α-MEM培養基培養細胞,每周換液2次,待細胞長滿培養皿后,0.05%胰蛋白酶消化并傳代,倒置相差顯微鏡下觀察細胞形態。
1.2.2 細胞擴增分析
SMSCs培養期間每隔3 d檢測細胞擴增情況。采用Proteinase K試劑盒提取細胞DNA,PicoGreen Assay測量細胞總DNA量,通過DNA量反映細胞擴增情況,繪制細胞擴增曲線。
1.2.3 SMSCs鑒定
① 流式細胞術:根據文獻[2]方法采用流式細胞儀鑒定原代貼壁細胞表面標志物CD44、CD90、CD105、CD14、CD34和CD45表達。
② 成軟骨分化:取20 μL第4代SMSCs,調整細胞濃度為2×107個/mL,接種于24孔板,貼壁3 h后加入成軟骨誘導培養液(含10 ng/mL TGF-β1、50 μg/ mL抗壞血酸、6.25 μg/mL胰島素、6.25 μg/ mL轉鐵蛋白、6.25 ng/mL亞硒酸、1.25 mg/mL BSA、5.35 mg/mL亞麻酸、100 nmol/L DEX的DMEM培養基),每2天更換培養基,維持TGF-β1濃度為10 ng/mL。培養14 d后PBS沖洗,- 20℃甲醇固定30 min,甲苯胺藍染色過夜,蒸餾水反復沖洗,以200 μL 6 mol/L鹽酸胍室溫下萃取,分光光度法測定波長630 nm處吸光度(A)值。
③ 成骨分化:如前步驟于24孔板內加入成骨誘導培養基(含100 nmol/L DEX、0.2 mmol/L ASA、10 mmol/L β-甘油磷酸鈉的DMEM培養基),培養21 d后加入0.2 nmol/L CaCl2,PBS沖洗后,4%多聚甲醛固定,2%茜素紅染色(pH4.2)3 min,蒸餾水沖洗,倒置相差顯微鏡下觀察。
④ 成脂分化:根據文獻[4]方法進行成脂分化鑒定。如前步驟于24孔板內加入成脂誘導培養基(含1 μmol/L DEX、0.5 mmol/L 3-異丁基-1-甲基黃嘌呤、100 μmol/L吲哚美辛、10 μg/mL胰島素的DMEM培養基)。誘導21 d后4%多聚甲醛固定1 h,0.5%油紅O染色2 h,倒置相差顯微鏡下觀察。
1.3 缺失實驗設計
首先使用缺失實驗方法,探索SMSCs成纖維軟骨分化的必要條件,再根據缺失實驗結果設計正交實驗。缺失實驗實驗條件包括:10 ng/ mL TGF-β3、500 ng/ mL BMP-2、100 nmol/L DEX、40 μg/mL 脯氨酸、50 μg/ mL ASA、100 μg/mL丙酮酸、6.25 μg/mL胰島素、6.25 μg/ mL轉鐵蛋白、6.25 μg/mL亞硒酸、1.25 mg/ mL BSA、5.35 mg/mL亞麻酸、10 ng/mL重組人TGF-β1、50 ng/mL bFGF、1.2%藻酸鹽溶液、循環間斷靜水壓(intermittent hydraulic pressure,IHP;0.5 kPa、1 Hz)、低氧環境(2%O2)、50 ng/mL BMP-7、10 ng/ mL IGF、10 ng/mL TGF-β2、50 ng/mL BMP-4、37℃、三維環境、5%CO2。上述條件中,三維環境采用小腸黏膜下層(small intestinal submucosa,SIS)支架,三維條件缺失時采用凝膠支架,實現單層培養。為明確溫度對SMSCs成纖維軟骨分化的影響,設立獨立對照實驗:采用隨機數字表法定義46.4、4.5、20.0℃作為溫度對照,并采用30~40℃連續0.5℃階梯變溫,探索溫度對SMSCs成纖維軟骨分化能力的調控作用。
取第4代SMSCs以2.5 ×105個/mL按上述實驗條件誘導,每組缺失1個條件(1~23組,見表 1),以加入所有誘導條件為陽性對照組(24組)。倒置相差顯微鏡下觀察各組細胞形態;培養14 d甲苯胺藍染色,倒置相差顯微鏡下觀察支架復合體形態及染色情況;并行半定量RT-PCR和免疫組織化學染色測定各組細胞是否表達SOX9、聚集蛋白聚糖(Aggrecan,AGN)、ColⅠ、ColⅡ以及ColⅨ。① 半定量RT-PCR:使用RNeasy Mini Kit試劑盒提取總RNA,使用SuperScriptⅢFirst-Strand Synthesis System Kit試劑盒逆轉錄得到cDNA,采用半定量RT-PCT對目的基因進行擴增,引物序列見表 2;在擴增進入平臺期前,檢測各孔A值,對基因濃度進行半定量分析,數據僅用于輔助分析成軟骨分化趨勢。② 免疫組織化學染色:采用免疫熒光法檢測高表達ColⅠ的細胞占軟骨細胞比例。封閉抗原后,采用鼠抗兔ColⅠ、ColⅡ一抗,連接了熒光素酶的馬抗鼠二抗,使用FITC和TRITC顯色。手工板計數,隨機取10個不同視野下10個大格內全部軟骨細胞作為總體,由于彈性軟骨細胞數量稀少,所以高表達ColⅠ的纖維軟骨細胞與高表達ColⅡ的透明軟骨細胞之和應等于軟骨細胞總體;高表達ColⅠ的纖維軟骨細胞計數與該總體比值視為纖維軟骨轉化率(N),以10個視野下各轉化單元N值的復合標準差作為實驗的總體標準差。實驗重復3次。
引入rate作為缺失實驗檢驗指標,rate定義為CD151+/CD44+細胞數與N值的乘積;再進一步結合細胞形態、甲苯胺藍染色、半定量RT-PCR檢測SOX9、AGN、ColⅠ、ColⅡ、ColⅨ基因表達來驗證rate值的信度。SMSCs成纖維軟骨分化后流式細胞計數和染色得到的N值必須與半定量RT-PCR檢測的基因表達趨勢一致,rate值才能用于統計分析,差異表達的結果不會出現在結果部分。為提高實驗信度,實驗平行3次,結果中報道的轉化值為最有代表性的實驗結果,而不是3次實驗的均值,存在交互作用的因子進一步行相關性分析,成直線正相關的兩個因子僅保留N值相對較高的因子。
1.4 正交實驗設計
根據缺失實驗篩選出調控條件:TGF-β1、BMP-2、DEX、脯氨酸、ASA、丙酮酸、胰島素+轉鐵蛋白+亞硒酸預混液(insulin + transferring + seleniousacid,ITS)、BSA、bFGF、IHP、BMP-7、IGF對SMSCs成纖維軟骨調節作用顯著(表 3),納入2水平正交實驗,各因子的水平參考文獻[1-5]獲得;溫度、CO2濃度和低氧環境存在交互作用,這些條件改變或缺失后SMSCs不能成纖維軟骨分化。其中,ITS預混溶液中各因子對SMSCs成纖維軟骨分化的影響存在直線正相關,此處將胰島素、轉鐵蛋白和亞硒酸作為一個因素設計為ITS。根據我們的實驗條件可進行單組<64孔的培養,選擇L60(212)正交設計能夠滿足實驗且最大限度保證結果準確性。使用SPSS18.0統計軟件設計正交實驗表頭(表 4),定義兩個水平值為水平1、水平2;根據表 1中的不同實驗條件對SMSCs行成纖維軟骨誘導。采用流式細胞儀計數,檢測CD151+/CD44+細胞并記錄N值;采用免疫組織化學染色,結合細胞形態、甲苯胺藍染色、半定量RT-PCR檢測SOX9、AGN、ColⅠ、ColⅡ、ColⅨ基因表達,進一步驗證結果。檢驗指標rate是CD151+/CD44+細胞與N值的乘積,計算方法同缺失實驗。
1.5 統計學方法
為防止不同個體來源SMSCs成纖維軟骨分化差異影響,采用來源于同一動物的SMSCs重復3 次實驗,取均值和標準差,并計算95%可信區間。來源于其余4只實驗動物的SMSCs作為對照,只有5組同時得到有意義的陽性結果,且結果趨勢一致,才能作為實驗結果,結果部分報道的N值是最典型的1只實驗動物的數據。缺失實驗采用方差分析,比較各組與標準條件組間的關系,排除非必要的調控因子;正交實驗結果采用直觀觀察和主體間方差分析方法,考慮部分因子間的1階交互作用,組間比較采用LSD和q檢驗,采用Ⅲ型平方和校正模型;檢驗水準α=0.05。
2 結果
2.1 SMSCs形態學觀察及鑒定
2.1.1 SMSCs形態學觀察及細胞擴增結果
原代細胞呈多邊形或星形;第1代細胞形態逐漸變成圓形和梭形;隨著傳代代次增加,細胞形態逐漸均一,呈“向日葵”樣分布,第3代細胞呈梭形。見圖 1。細胞倍增時間為28 h,細胞擴增曲線見圖 2。
圖1
SMSCs形態學觀察(倒置相差顯微鏡×40) ? 原代細胞 ? 第3代細胞 ? ?2.1.2 SMSCs鑒定
流式細胞儀檢測示SMSCs表面標志CD44、CD90、CD105呈陽性表達,CD14、CD34、CD45呈陰性表達。多向分化鑒定示,成骨誘導21 d茜素紅染色、成軟骨誘導14 d甲苯胺藍染色及成脂誘導21 d油紅O染色均呈陽性(圖 3)。
2.2 SMSCs體外成軟骨分化觀察
按照正交實驗培養條件誘導3 d后,細胞聚集成小團狀,其趨化因子不清楚,細胞沿支架間隙爬行生長;支架體積在培養5 d倍增;14 d細胞-支架復合物甲苯胺藍染色呈陽性(圖 4)。軟骨細胞免疫組織化學染色示ColⅠ、ColⅡ呈陽性染色(圖 5)。半定量RT-PCR檢測示,誘導后細胞表達SOX9、AGN、ColⅠ、ColⅡ、ColⅨ基因(圖 6),各組基因表達不同,誤差線表示50%可信區間。
2.3 缺失實驗結果
缺失實驗結果顯示,TGF-β1、BMP-2、DEX、脯氨酸、抗壞血酸、丙酮酸、ITS、BSA、bFGF、IHP、BPM-7、IGF對SMSCs成纖維軟骨分化調控作用顯著(表 5),且各因子間不存在交互作用,作為獨立因子參與正交實驗。溫度降低、升高,高氧環境以及使用凝膠限制細胞接觸的狀態下,細胞誘導分化明顯延遲。來源于同一細胞株的SMSCs,限制細胞接觸后成纖維軟骨分化能力不同;溫度對SMSCs成纖維軟骨分化的影響并非成線性,在35.5~38.5℃范圍內,成纖維軟骨分化能力無顯著差異,當溫度高于或低于此范圍時,SMSCs成纖維軟骨分化能力下降。因此,溫度、氧環境以及細胞間接觸是SMSCs成纖維軟骨分化必不可少的條件,但條件本身在閾值范圍內改變時影響并不顯著,超過閾值后SMSCs幾乎不能成纖維軟骨分化,因此不作為正交實驗因子。亞麻酸缺失對成纖維軟骨分化影響不顯著,且與其他因子不存在交互作用,為非必要調控因素;方差分析顯示TGF-β1、TGF-β3與 TGF-β2存在交互作用,成線性正相關,且不與其他因子交互,TGF-β1、TGF-β2與TGF-β3同時缺失,SMSCs不能成纖維軟骨分化,缺失TGF-β1時SMSCs成纖維軟骨分化能力較強,但纖維軟骨細胞分化率僅為1.18%,而缺失TGF-β3、TGF-β2時為61.89%和75.13%,因此保留TGF-β1作為調控因子。藻酸鹽缺失狀態下,SMSCs成纖維軟骨分化率為62.64%,與陽性對照組間比較差異無統計學意義(P>0.05),且與各指標間無交互作用,為非必要調控因素。
2.4 正交實驗結果
正交實驗結果見表 6,直觀分析顯示,按轉化率降序擴展排序后,取轉化率中位數為分界線區組觀察,第1因子TGF-β1對轉化率的作用最明顯,TGF-β1高濃度組轉化率均>30%、低濃度組轉化率<30%;其余分界線兩側的水平值加和均為45,直觀條件下無法判斷其轉化率差異。取轉化率1/4位數區組觀察,見第11因子BMP-7第1區組轉化率最高,水平值加和顯示第1區組為30,第2、3、4區組(轉化率依次降低)水平值加和分別為15、23、22,說明BMP-7采用水平2有利于得到更高的纖維軟骨轉化率;其余第1區組水平值加和高于22.5的因子有第3、5、7、9、12因子,第4區組的水平值加和均低于22.5,說明采用本正交實驗表頭設計的正交實驗,各因子與轉化率之間有較好的相關性。
正交實驗方差分析采用直觀觀察和主體間方差分析的方法,結果見表 6。校正模型P=0.000,表明該模型能夠滿足實驗設計;截距P=0.000,表明各因子對因變量影響差異不完全相同。TGF-β1、ASA、bFGF、IGF、BMP-7等因子對因變量調控作用較其他因子顯著,差異有統計學意義;即改變以上因子濃度,SMSCs成纖維軟骨分化能力顯著降低,上述因子是SMSCs成纖維軟骨分化最重要的調節因素,與直觀觀察結果相似。
3 討論
本研究結果表明,SMSCs成纖維軟骨分化過程中,TGF-β1、ASA、bFGF、IGF 4個因子劑量的改變顯著影響SMSCs成纖維軟骨分化作用,通過合理調整上述因子濃度,可顯著提高成纖維軟骨細胞分化轉化率;但由于檢索資源限制,我們的實驗設計不可能覆蓋所有敏感因子,這4個因子并不是SMSCs成纖維軟骨分化唯一調控因素。TGF-β是公認的SMSCs向軟骨細胞分化最關鍵的啟動因素[1-2],但對不同亞型的TGF-β促MSCs成纖維軟骨分化作用存在爭議[3-5]。Kim等[5]指出瞬間暴露在高劑量TGF-β3環境中,MSCs功能和表型被誘導變異,并能在體內增強軟骨修復能力。Vanneaux等[3]認為TGF-β受體Ⅱ在TGF-β1介導的MSCs向軟骨細胞分化過程中至關重要,TGF-β1是介導SMSCs成纖維軟骨分化的最重要因子。我們的實驗表明,不同亞型的TGF-β誘導MSCs成纖維軟骨分化能力不同,TGF-β1調節作用最強。胰島素是MSCs成纖維軟骨分化中必要的調節因子[6-7],但本實驗結果表明胰島素對SMSCs成纖維軟骨分化的調節作用較弱,指數級的劑量差異仍不能引起纖維軟骨轉化率的差異,且與TGF和亞硒酸之間存在線性交互作用,不能作為獨立的調節因子。
BMP亦是重要的纖維軟骨生成調節因子[8],許多文獻定義BMP為促進劑[9],但是對各亞型BMP作用存在爭議[10-11]。我們的實驗表明BMP-2促SMSCs成纖維軟骨分化能力最強,已經明確BMP-2調節通路是:BMP-2→TGF-β受體Ⅱ→Smad 1、5、8通路,可能由于受體的飽和作用,BMP-2調控存在天花板效應[12],高于調控濃度上限的BMP-2不會增加SMSCs成纖維軟骨分化能力,我們的實驗結果表明500 ng/ mL BMP-2與100 ng/mL BMP-2間差異不顯著,但是與缺失組對照,其促軟骨生成能力顯著提高。此外,BMP-2促MSCs成纖維軟骨分化作用機制尚存在爭議。壓力作為獨立調控因素能夠通過調節內源性BMP-2表達來調控MSCs成纖維軟骨分化過程;同時,壓力因素與BMP-2存在交互作用,Valhmu等[13]實驗證實壓力具有促進MSCs成纖維軟骨分化和促進軟骨細胞分泌AGN的作用。Wong等[14]研究也證實了壓力能夠促進軟骨細胞基質分泌。Feng等[15-16]認為鼠類BMP-2、4的啟動子區域存在與壓力應答共存的區域,這可能是壓力調控的基因基礎。Rui等[17]、Lui等[18]、Ando等[19]均認為細胞外基質的破壞和重構與該過程密切相關。Chikazu等[20]提出了BMP-2→COX2→前列腺素E2(prostaglandin E2,PGE2)(Cbfa)→骨/軟骨形成的調控模型,認為PGF2存在對BMP-2的抑制作用,Cbfa基因的高表達促進成骨作用,抑制成軟骨作用,可能與BMP-2濃度的天花板效應相關。Siddhivarn等[21]提出的Δ12-PGF J2/過氧化物酶體增殖物激活受體-γ2/BMP-2通路可能是壓力與BMP-2相關的調節通路。
ASA在MSCs成骨分化中的作用及調控通路已被廣泛研究[22-23],在成纖維軟骨分化過程中,ASA是MSCs重要細胞因子,但其在成纖維軟骨分化過程中的調控作用研究較少[24]。關于ASA和SMSCs間相互作用的報道,在筆者委托的查新報告中未見收獲。我們的實驗表明,ASA在SMSCs成纖維軟骨分化過程中可作為單獨的調控因子,且與其他因子不存在交互作用。bFGF促進MSCs成軟骨分化的作用是明確的[25],其調節通路主要通過抑制α-平滑肌動蛋白的表達,維持成軟骨必要的三維形態,從而促進MSCs成軟骨分化過程。Kim等[26]研究證實,10 ng/ mL bFGF在成纖維軟骨前的預處理能顯著促進SMSCs成纖維軟骨分化作用;值得注意的是,同組研究者在2009年卻發布了截然相反的報道,Lee等[27]認為加入20 ng/mL bFGF和血清后,成纖維軟骨分化過程受到顯著抑制;這一因子的調節通路至今尚未得到深入研究。我們的實驗結果表明,50 ng/ mL bFGF較空白對照組顯著增強了SMSCs成纖維軟骨分化能力,成透明軟骨能力顯著下降,但在加入高濃度(200 ng/ mL)bFGF后,SMSCs成纖維軟骨和透明軟骨能力均顯著下降;針對bFGF可能調節通路的下游因子實驗,我們在5組動物實驗中得到矛盾的結果,故未在本文報道,目前我們仍不能確定該因子的作用機制,將在后續研究中進一步探討。
其他作用因子對MSCs成纖維軟骨分化的作用存在爭議,如DEX作為獨立調節因子作用不穩定,Nogami等[9]報道DEX能夠增強人MSCs成纖維軟骨分化過程,但Liu等[28]報道認為無論是否添加DEX,TGF-β2均能很好誘導成軟骨,我們的研究同樣表明,DEX作為獨立因子,對SMSCs成纖維軟骨調控能力不顯著,少量DEX(<100 μg/ mL)能促進SMSCs成纖維軟骨分化,但大劑量DEX(>1 000 μg/ mL)對SMSCs成脂分化表現出更強的促進作用。此外,我們觀察到擴增階段培養基中糖和血清的濃度對成軟骨分化存在調控作用,目前未見相關報道,這一作用有待深入研究。
綜上述,本文采用正交實驗方法,探索了SMSCs成纖維軟骨分化條件,找到了效率相對較高的SMSCs成纖維軟骨分化實驗條件,但細胞生長是一動態變化過程,許多條件的時間相關性還需深入探索。