引用本文: 張臣鳴, 李宇, 張玉強, 曹宇, 龔超, 王晨亮, 王偉. 周圍神經損傷后斷端神經組織微小 RNA-221 和 PTEN 表達變化. 中國修復重建外科雜志, 2019, 33(9): 1162-1168. doi: 10.7507/1002-1892.201903122 復制
版權信息: ?四川大學華西醫院華西期刊社《中國修復重建外科雜志》版權所有,未經授權不得轉載、改編
周圍神經損傷(peripheral nerve injury,PNI)后經典的修復過程是神經遠側斷端瓦勒變性溶解消失,重新由近側斷端向遠處生長出整個遠側神經[1-4]。大量研究證明,磷酸酶與張力蛋白同源物(phosphatase and tension protein homologue,PTEN)在 PNI 后的修復中起重要作用,其蛋白表達量降低可以促進軸突及髓鞘再生[5-7]。近年來,微小 RNA(microRNA,miRNA)因在各種生物學功能中的調節作用引起了極大關注。有研究證實,在多種腫瘤細胞中,miR-221 對 PTEN 具有調控作用[8-10],其通過與 PTEN mRNA 3′端非翻譯區(3′UTR)互補結合來抑制 PTEN 磷酸水解酶表達,從而減弱 PTEN 在控制細胞增殖、凋亡、轉移與侵襲中的效能[11]。然而,miR-221/PTEN 在 PNI 后修復中是否存在相關性未見報道。此外,PNI 后修復受到神經遠側斷端發出的信號調控[12-13],這種不對稱修復現象使軸突再生由神經近側斷端向遠側斷端生長[14],miR-221/PTEN 在近、遠側斷端表達變化是否存在差異尚未明確。本實驗中,我們旨在研究 miR-221 與 PTEN 在 PNI 后修復中的表達及調控關系,比較近、遠側斷端中二者表達差異,從而為臨床診治 PNI 提供新靶點。
1 材料與方法
1.1 實驗動物、細胞及主要試劑、儀器
SPF 級雄性 SD 大鼠 96 只,體質量(200±20)g,由錦州醫科大學實驗動物中心提供,于光照和黑暗每 12 小時交替環境中標準飼養。293T 細胞(ATCC 細胞庫,美國)。
FBS(Serapro 公司,美國);DMEM 培養基、青/鏈霉素、胰蛋白酶(HyClone,美國);Opti-MEM 培養基 (GIBCO 公司,美國);兔抗鼠 PTEN 多克隆抗體(北京博奧森生物技術有限公司);兔抗鼠 GAPDH 多克隆抗體、辣根過氧化物酶標記的山羊抗兔 IgG 二抗(Earthox 公司,美國);Alexa Fluor 647-結合羊抗兔二抗(Abcam 公司,英國);牛血清白蛋白(Sigma 公司,美國);蛋白提取試劑盒、BCA 蛋白定量試劑盒、SDS-PAGE 凝膠試劑盒、ECL 化學發光試劑盒(上海碧云天生物技術有限公司);蛋白 Marker、GeneJETTMRNA 純化試劑盒(Thermo Fisher 公司,美國);GoScriptTM Reverse Transcription System 逆轉錄試劑盒、GoTaq? qPCR Master Mix 試劑盒、Dual-Luciferase? Reporter Assay System 試劑盒、pmirGLO Dual-Luciferase? miRNA Target Expression Vector(Promega 公司,美國);miR-221 模擬物(蘇州吉瑪基因股份有限公司);LipofectamineTM 2000(Invitrogen 公司,美國);QuikChange? Lightning Site-Directed Mutagenesis Kits 試劑盒(Stratagene 公司,美國);Gluta 固定液(Solarbio 公司,美國)。
顯微外科手術器械(泗洪金睛醫療器械有限公司);病理切片機、超薄切片機(Leica 公司,德國);正置熒光顯微鏡(Nikon 公司,日本);紫外分光光度計(Tecan 公司,美國);全自動電泳儀、化學發光凝膠成像系統(Bio-Rad 公司,美國);ABI stepone plus 實時熒光定量 PCR 儀(Applied Biosystems 公司,美國);透射電鏡(Hitachi 公司,日本)。
1.2 細胞培養
取 293T 細胞采用含 10%FBS 的 DMEM 培養基培養,置于 37 ℃、5%CO2 細胞孵育箱中培養。
1.3 大鼠坐骨神經損傷模型建立
取 96 只 SD 大鼠環境適應處理 12 h 后,100 g/L 水合氯醛(350 mg/kg)腹腔注射麻醉;麻醉后靜置動物約 3 min,備皮后于右下肢股骨中段下緣 5 mm 處作平行于股骨的切口約 5 mm[15],組織鉗鈍性分離筋膜及肌間隙后,暴露坐骨神經,用顯微外科剪離斷坐骨神經,觀察動物神經反射情況。逐層縫合后注射阿米卡星 0.1 mL 預防感染,切口包扎。待麻醉復蘇后,觀察動物運動恢復情況。分別于術后 0(術后即刻)、1、4、7 d,以麻醉后頸椎脫臼法分別處死 24 只大鼠,無菌條件下取坐骨神經近、遠側斷端組織各 5 mm 進行以下觀測。
1.4 觀測指標
1.4.1 實時熒光定量PCR檢測miR-221基因表達
于術后各時間點分別取 6 只大鼠坐骨神經近、遠側斷端組織,液氮速凍后研磨成粉,用 RNA 純化試劑盒過柱法提取組織總 RNA,經紫外分光光度計檢測,測定吸光度(A)值(A260/A280 值在 1.9~2.2 之間用于 cDNA 合成)。按照逆轉錄反應體系進行逆轉錄,所得 cDNA 在實時熒光定量反應體系下擴增,擴增 40 個循環。引物序列:miR-221 上游 5′-ATGCTCTAAGCTACATTGTCTGC-3′,下游 5′-TATGGTTGTTCTGCTCTCTGTCTC-3′;內參照 U6 上游 5′-CGCTTCGGCAGCACATATAC-3′,下游 5′-TTCACGAATTTGCGTGTCATC-3′。采用 2?ΔΔCt法計算 miR-221 相對表達量,實驗重復 3 次,取均值。
1.4.2 Western blot檢測PTEN蛋白表達
于術后各時間點分別取 6 只大鼠坐骨神經近、遠側斷端組織,于冰上按 1 mg 神經組織∶20 μL RIPA 裂解液進行裂解,4 ℃ 冰浴勻漿 30 min 后,以離心半徑 3 cm、12 000 r/min 離心 30 min,取上清液即為所需蛋白液。BCA 法蛋白定量后調整至同等濃度,每孔上樣 30 μg,經 10%SDS-PAGE 凝膠電泳后轉移至聚偏二氟乙烯膜,1% 牛血清白蛋白封閉 1 h;加入 TBST 稀釋兔抗鼠 PTEN 多克隆抗體(1︰1 000)、兔抗鼠 GAPDH 多克隆抗體(1︰3 000),4 ℃ 孵育過夜;TBST 洗膜 3 次,加入辣根過氧化物酶標記的羊抗兔 IgG 二抗(1︰10 000)室溫孵育 2 h;TBST 洗膜 3 次,在暗室內行化學發光劑 ECL 反應,化學發光凝膠成像系統取像并測定條帶灰度值。以 PTEN 蛋白條帶和 GAPDH 內參條帶灰度值的比值作為目的蛋白的相對表達量。
1.4.3 雙熒光素酶報告基因實驗
通過 Targetscan、miRDB、Miranda 和 PicTar 等生物信息學軟件預測 miR-221 靶向結合 PTEN 3′UTR 的位點進行分析。利用 PCR 擴增包含 miR-221 結合位點的 PTEN 3′UTR 序列,將其克隆到 pmirGLO 報告載體,構建野生型 pmirGLO-PTEN-WT 報告載體;用單點突變試劑盒突變 pmirGLO-PTEN-WT 上的 miR-221 結合位點,構建突變型 pmirGLO-PTEN-Mut 報告載體。取對數生長期 293T 細胞,以 1.5×104個/孔接種于 96 孔板中。將 Opti-MEM 培養基稀釋的野生型和突變型報告載體分別與 miR-221 模擬物(miR-221 mimic)或 miR-陰性對照(miR-NC)混合,混合后加入 LipofectamineTM 2000 試劑。將所得混合液分別與 Opti-MEM 培養基以 1∶1 比例加入 293T 細胞,其轉染濃度報告載體為每孔 250 ng,miR-221 mimic 及 NC 為每孔 50 nmol/L 孔。培養 6 h 后再加 100 μL Opti-MEM 培養基,48 h 后用 Dual-Luciferase? Reporter Assay System 試劑盒檢測各組細胞螢火蟲熒光酶熒光值(M1)及海腎熒光素酶熒光值(M2),以 M1/M2 比值作為相對熒光值。
1.4.4 免疫熒光染色觀察PTEN蛋白表達
于術后各時間點分別取 6 只大鼠坐骨神經近、遠側斷端組織,于 4% 多聚甲醛固定 24 h,行脫水、石蠟包埋、切片等處理,常規行抗原修復、山羊血清封閉后,滴加兔抗鼠 PTEN 多克隆抗體(1∶400)4 ℃ 孵育過夜;置于搖床 PBS 洗滌 3 次,Alexa Fluor 647-結合羊抗兔二抗(1∶500)避光室溫孵育 30 min;DAPI 染液處理后,洗滌并用抗熒光淬滅封片劑封片,正置熒光顯微鏡下觀察 PTEN 蛋白在神經斷端橫截面上的分布。取 100 倍鏡下單個神經橫截面視野,應用 Image-Pro Plus 6.0 軟件分析其平均吸光度(IA)值作為 PTEN 蛋白相對表達量。
1.4.5 透射電鏡觀察
于術后各時間點分別取 6 只大鼠坐骨神經近、遠側斷端組織,取 1 mm×1 mm×1 mm 神經組織,Gluta 固定液固定 1~4 h,再用 1% 鋨酸-0.1 mol/L PBS 室溫固定 2 h,脫水、滲透、包埋后制作厚 60 nm 超薄切片,經鈾鉛雙染色后透射電鏡觀察神經軸突、髓鞘等超微結構的形態學變化。
1.5 統計學方法
采用 SPSS19.0 統計軟件進行分析。數據以均數±標準差表示,同側斷端各時間點間比較采用單因素方差分析,兩兩比較采用 Bonferroni 法;左、右側斷端同一時間點間比較采用獨立樣本 t 檢驗;雙熒光素酶報告基因實驗組間比較采用 Dunnett’s t 檢驗;檢驗水準 α=0.05。
2 結果
2.1 實時熒光定量PCR檢測miR-221表達
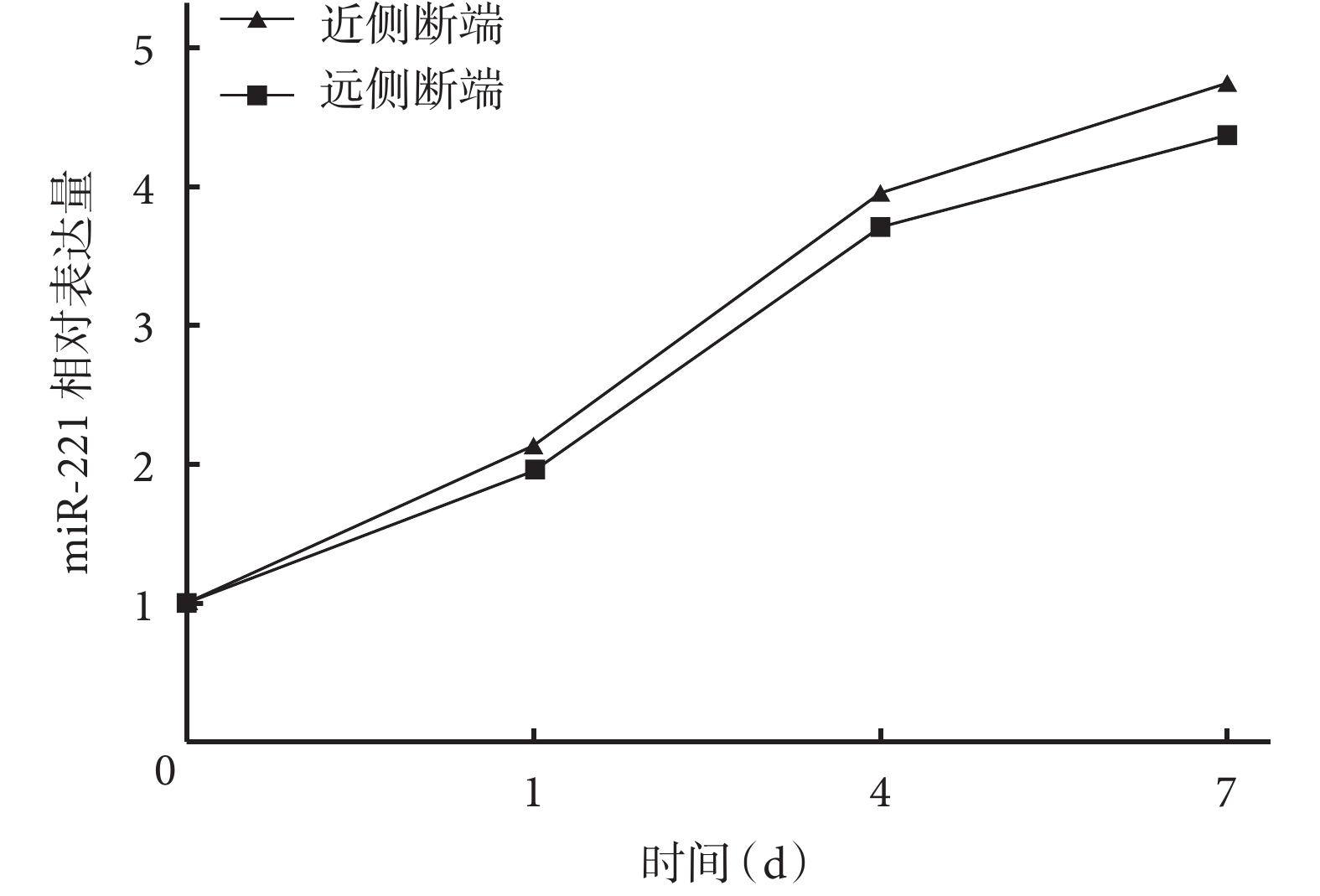
隨時間延長,大鼠坐骨神經近、遠側斷端組織中 miR-221 相對表達量均逐漸增加,術后各時間點間比較差異均有統計學意義(P<0.05)。術后 1、4、7 d 近側斷端組織中 miR-221 相對表達量均顯著高于遠側斷端,差異有統計學意義(P<0.05)。見圖1。
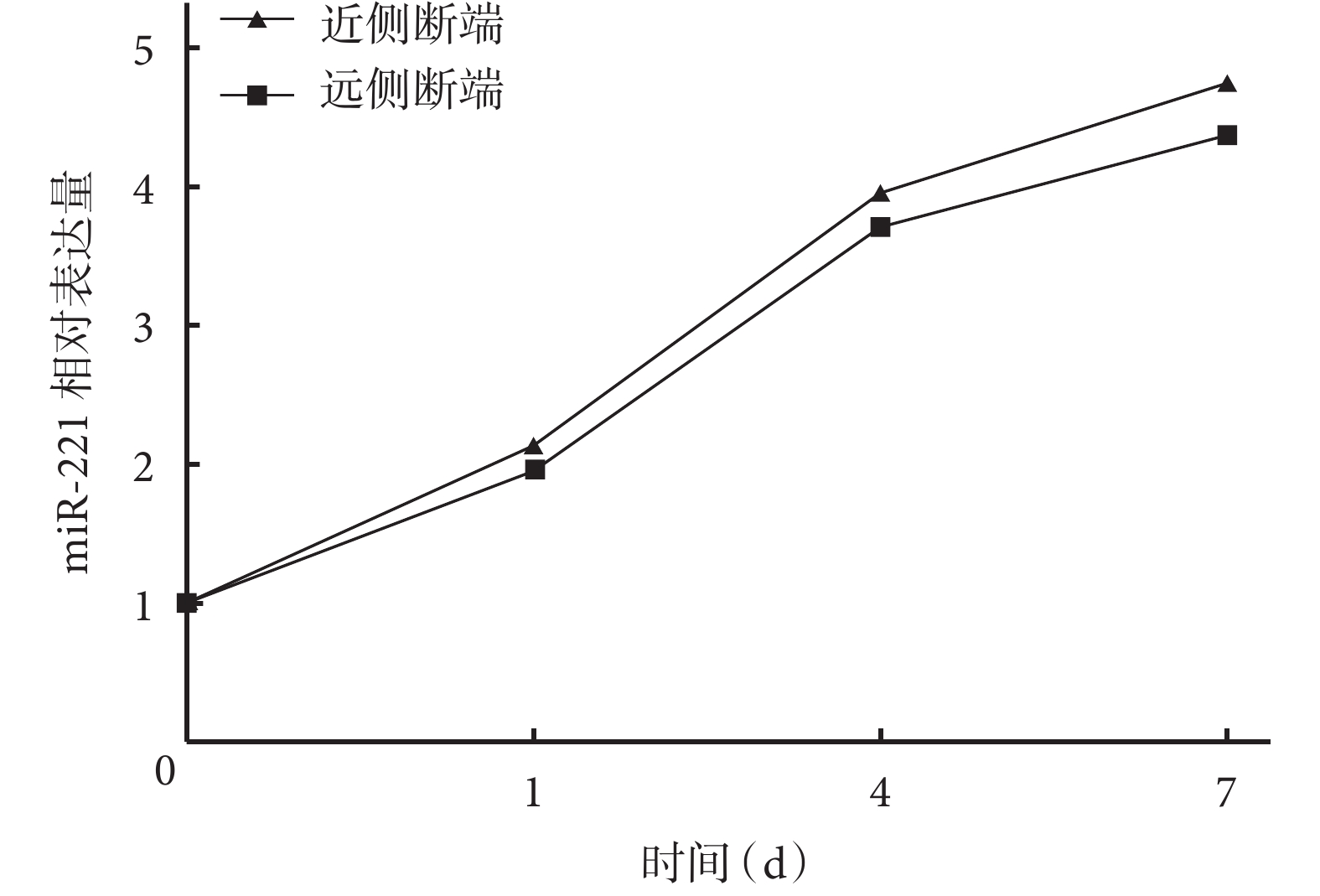 圖1
實時熒光定量PCR檢測術后各時間點大鼠坐骨神經 近、遠側斷端組織中miR-221相對表達量
Figure1.
The relative expression of miR-221 in the proximal and distal stumps at different time points after operation by real-time fluorescent quantitative PCR
圖1
實時熒光定量PCR檢測術后各時間點大鼠坐骨神經 近、遠側斷端組織中miR-221相對表達量
Figure1.
The relative expression of miR-221 in the proximal and distal stumps at different time points after operation by real-time fluorescent quantitative PCR
2.2 Western blot檢測PTEN蛋白表達
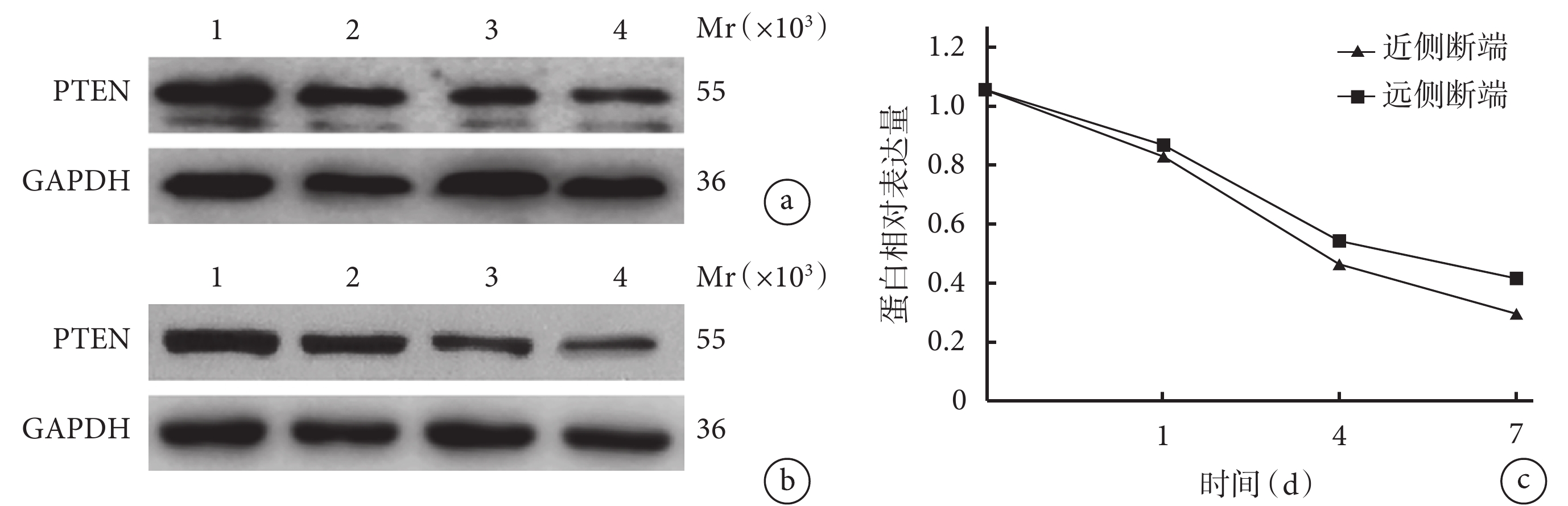
隨時間延長,大鼠坐骨神經近、遠側斷端組織中 PTEN 蛋白相對表達量均逐漸減少,術后各時間點間比較差異均有統計學意義(P<0.05)。術后 1、4、7 d 近側斷端組織中 PTEN 蛋白相對表達量均顯著低于遠側斷端,差異有統計學意義(P<0.05)。見圖2。
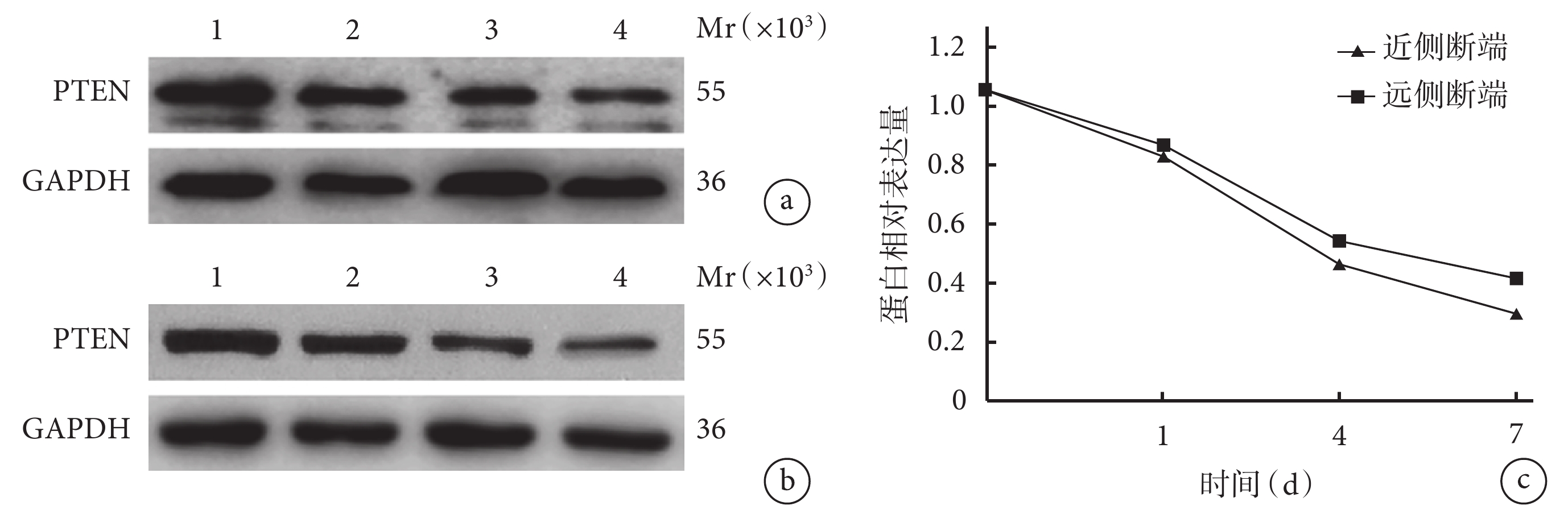 圖2
Western blot檢測術后各時間點大鼠坐骨神經近、遠側斷端組織中PTEN蛋白表達
圖2
Western blot檢測術后各時間點大鼠坐骨神經近、遠側斷端組織中PTEN蛋白表達
Mr:相對分子質量 1:0 d 2:1 d 3:4 d 4:7 d a. 近側斷端蛋白條帶圖;b. 遠側斷端蛋白條帶圖;c. PTEN 蛋白相對表達量
Figure2. The expression of PTEN protein in the proximal and distal stumps at different time points after operation detected by Western blotMr: Relative molecular mass 1: 0 day 2: 1 day 3: 4 days 4: 7 days; a. Protein bands of the proximal stump; b. Protein bands of the distal stump; c. Relative expression of PTEN protein
2.3 雙熒光素酶報告基因實驗
生物信息預測結果表明,miR-221 序列上存在 PTEN 的連續結合序列,見表1。通過構建野生型和突變型報告載體,雙熒光素酶報告基因實驗發現,與 miR-NC(1.003±0.009)相比,miR-221 mimic 與野生型報告載體共轉染時顯著降低了熒光素酶相對活性(0.402±0.045),差異有統計學意義(t=15.78,P=0.00);而與突變型報告載體共轉染時恢復了熒光素酶相對活性(0.972±0.021),差異無統計學意義(t=1.86,P=0.14)。
2.4 免疫熒光染色觀察PTEN蛋白表達
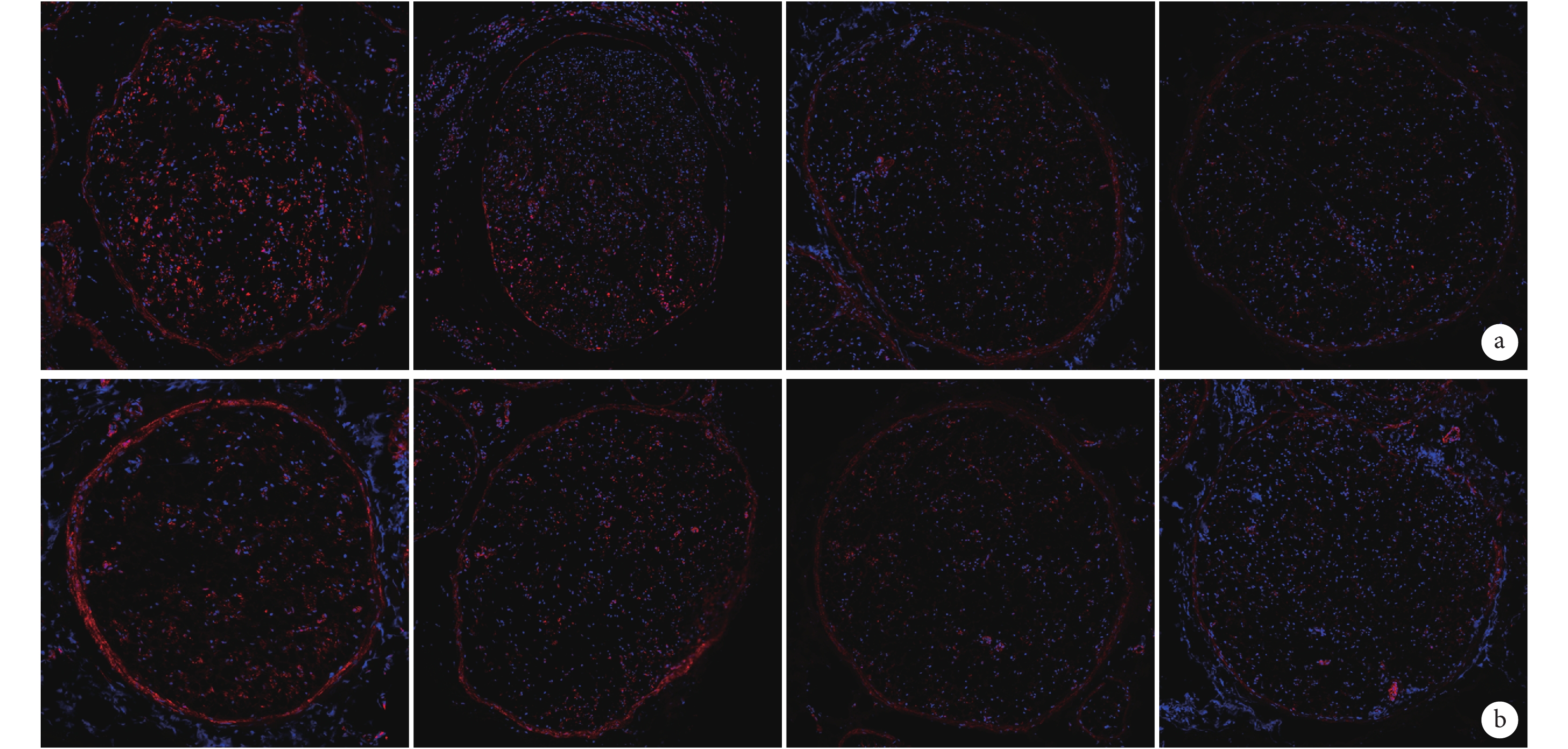
熒光顯微鏡觀察示,PTEN 蛋白主要表達于細胞質,部分細胞核中也有少許表達。隨神經損傷后修復時間的延長,PTEN 陽性表達明顯減少,其亞細胞定位無明顯變化。見圖3。
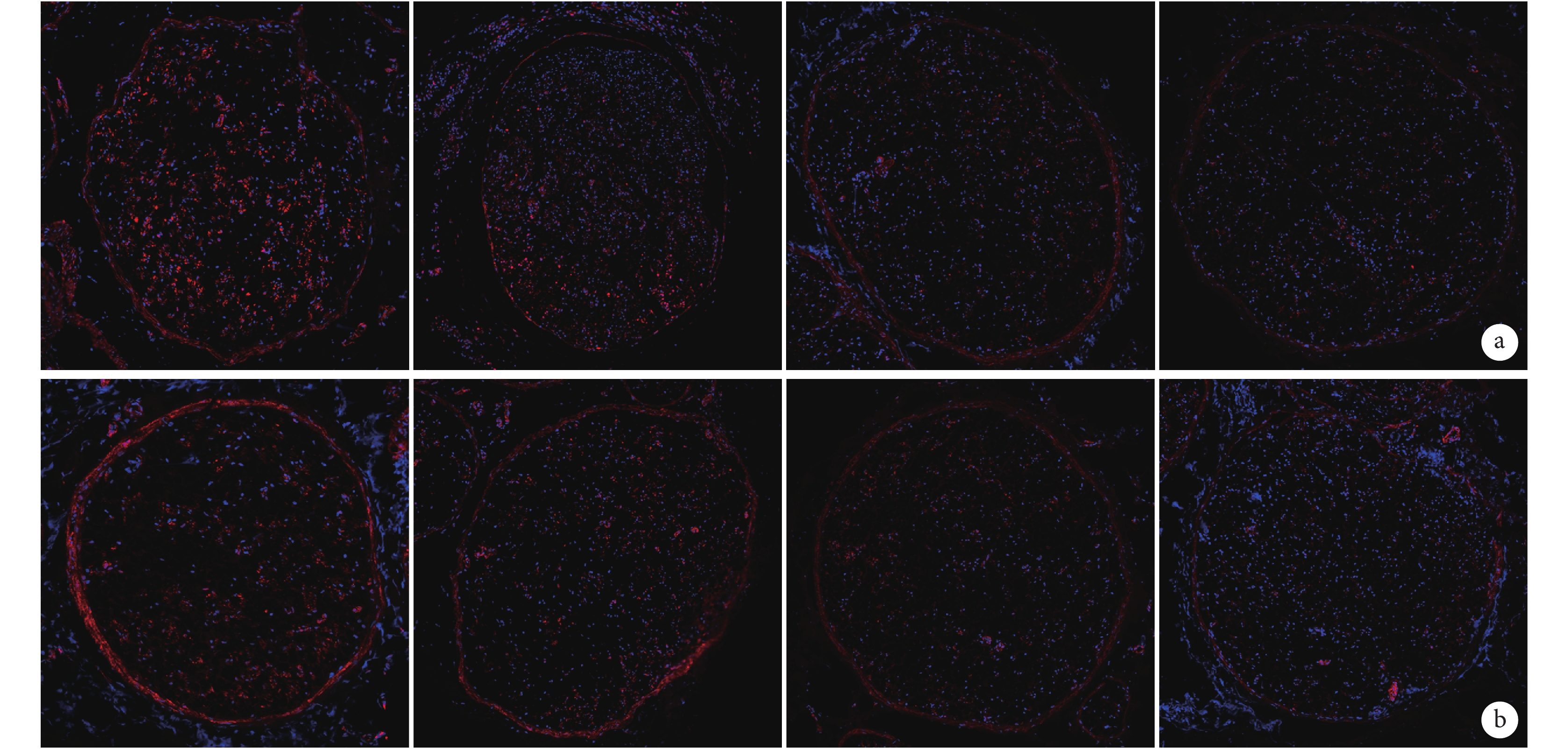 圖3
免疫熒光染色觀察術后各時間點坐骨神經近、遠側斷端組織中PTEN蛋白表達(熒光顯微鏡×100)
圖3
免疫熒光染色觀察術后各時間點坐骨神經近、遠側斷端組織中PTEN蛋白表達(熒光顯微鏡×100)
從左至右依次為術后 0、1、4、7 d a. 近側斷端;b. 遠側斷端
Figure3. Immunofluorescence staining observation of the expression of PTEN protein in the proximal and distal stumps at different time points after operation (Fluorescence microscopy×100)From left to right for postoperative views at 0, 1, 4, and 7 days, respectively a. Proximal stump; b. Distal stump
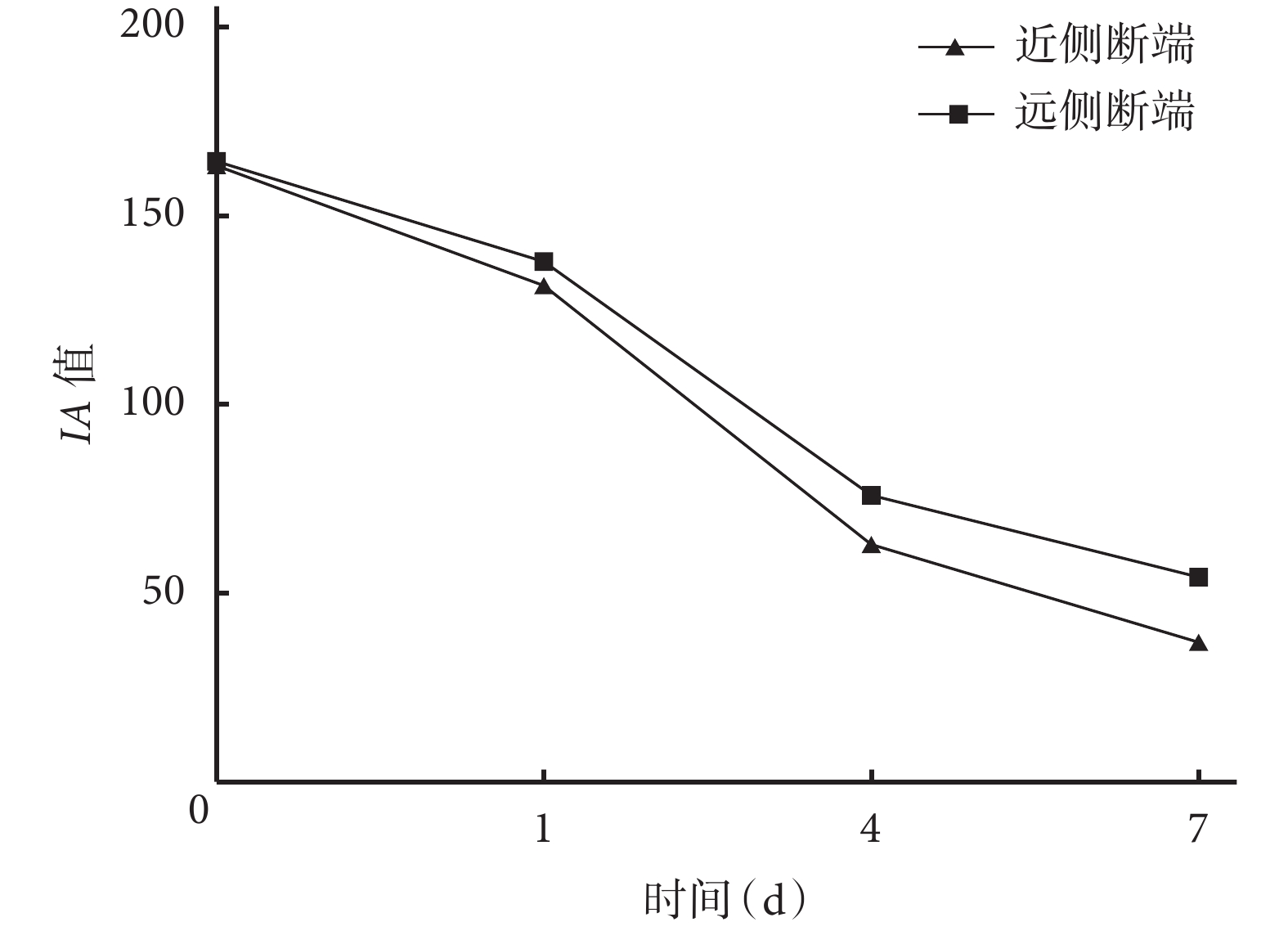
定量檢測示隨時間延長,大鼠坐骨神經近、遠側斷端組織 IA 值均逐漸減小,術后各時間點間比較差異均有統計學意義(P<0.05)。術后 1、4、7 d 遠側斷端組織 IA 值均顯著高于近側斷端,差異有統計學意義(P<0.05)。見圖4。
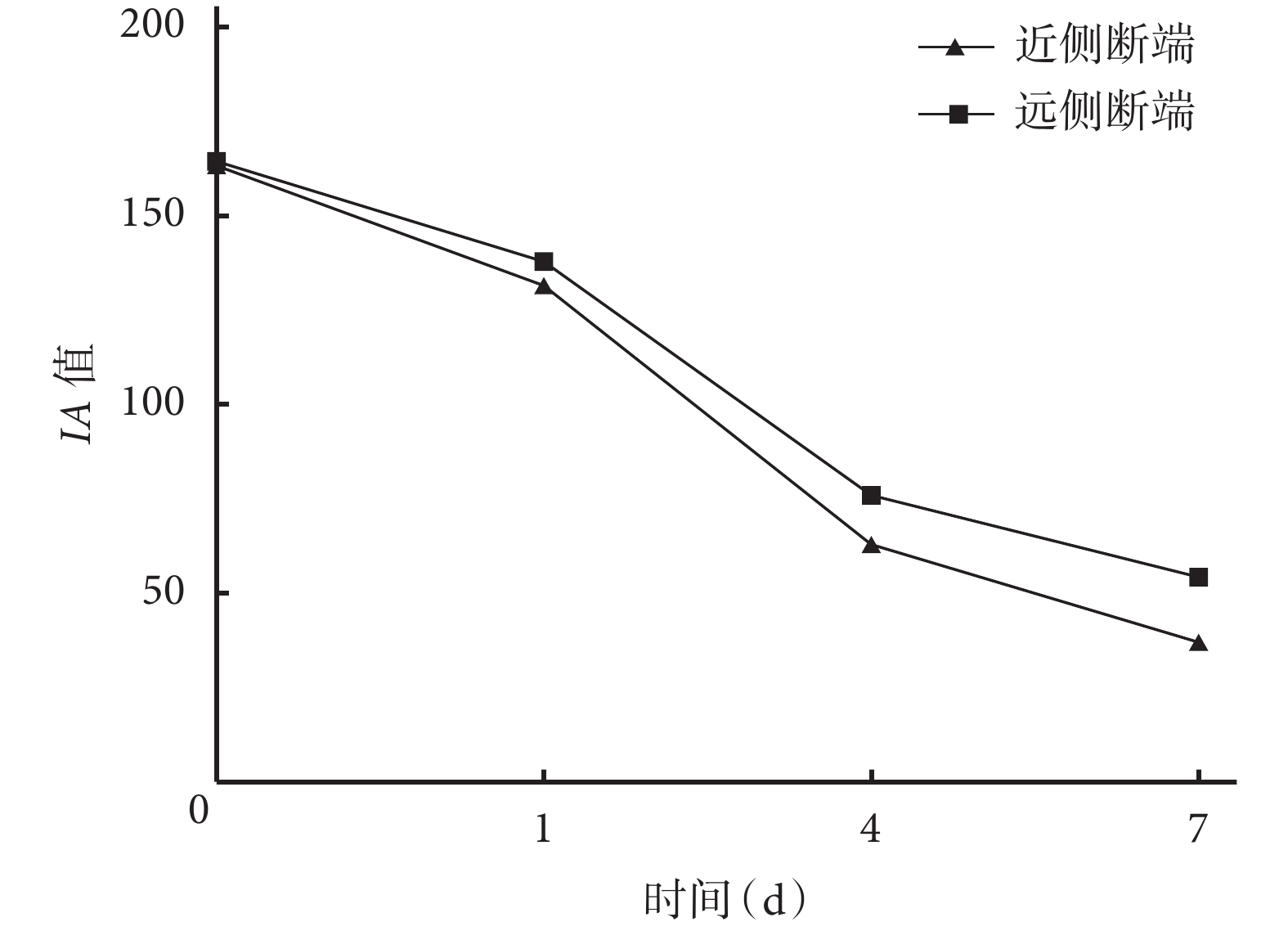 圖4
免疫熒光染色檢測術后各時間點坐骨神經近、遠側斷端 組織IA值
Figure4.
Detection of the IA value in the proximal and distal stumps at different time points after operation by immuno fluorescence staining
圖4
免疫熒光染色檢測術后各時間點坐骨神經近、遠側斷端 組織IA值
Figure4.
Detection of the IA value in the proximal and distal stumps at different time points after operation by immuno fluorescence staining
2.5 透射電鏡觀察
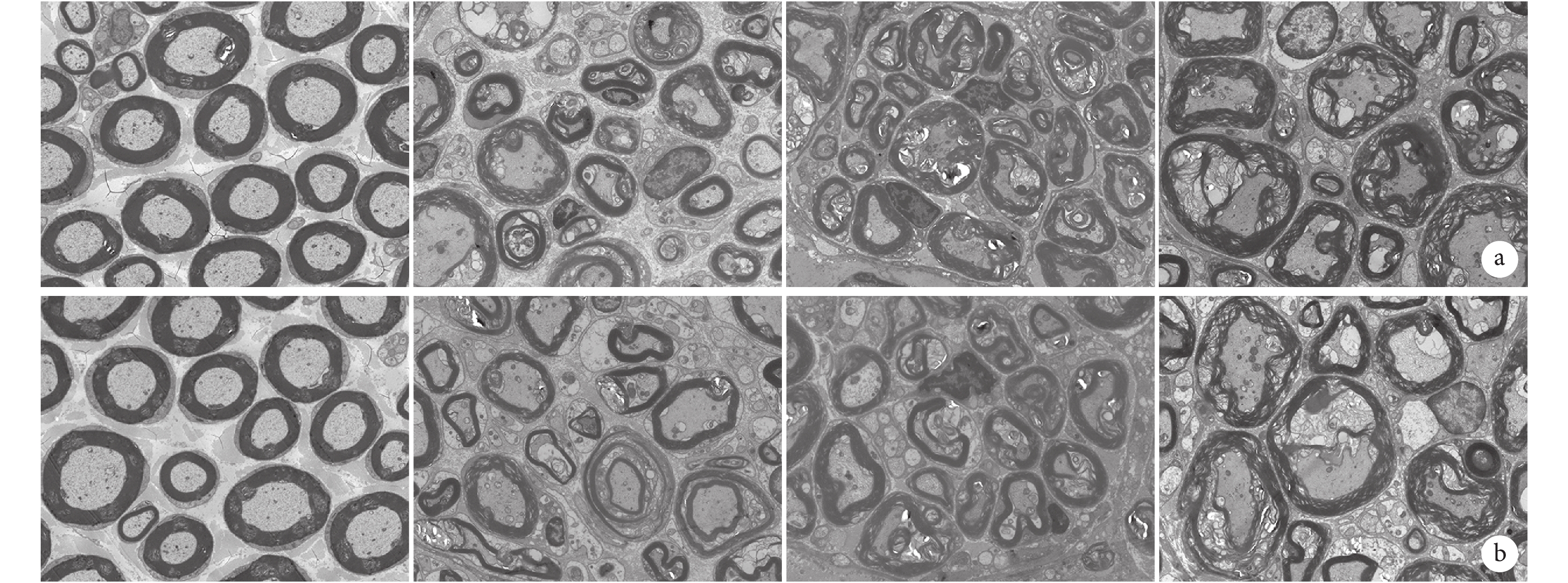
術后 0 d,近、遠側斷端正常軸突、髓鞘等超微結構形態規整、結構清晰,無病理改變。術后 1 d,近、遠側斷端雪旺細胞增殖及潰變均不明顯,髓鞘整體形態結構與術后 0 d 接近,但髓鞘板層略有疏松,鞘內軸漿顆粒消失。術后 4 d,雪旺細胞較 0 d 時明顯增殖旺盛,呈較明顯神經潰變,近、遠側斷端可見大量微管及微絲崩解,線粒體腫脹,髓鞘開始破壞,髓鞘內板層結構模糊,間隙增寬、松散,形態不規則;與遠側斷端相比,近側斷端雪旺細胞分裂、增殖更明顯,且軸突、髓鞘破壞程度更小。術后 7 d,已無完整髓鞘結構,雪旺細胞遷移引導軸突再生,近、遠側斷端神經潰變達到高峰,且遠側斷端神經潰變較近側斷端更徹底;近側斷端可見少數再生軸突已被雪旺細胞包繞,以及少數新生神經纖維及未成熟的新生髓鞘。見圖5。
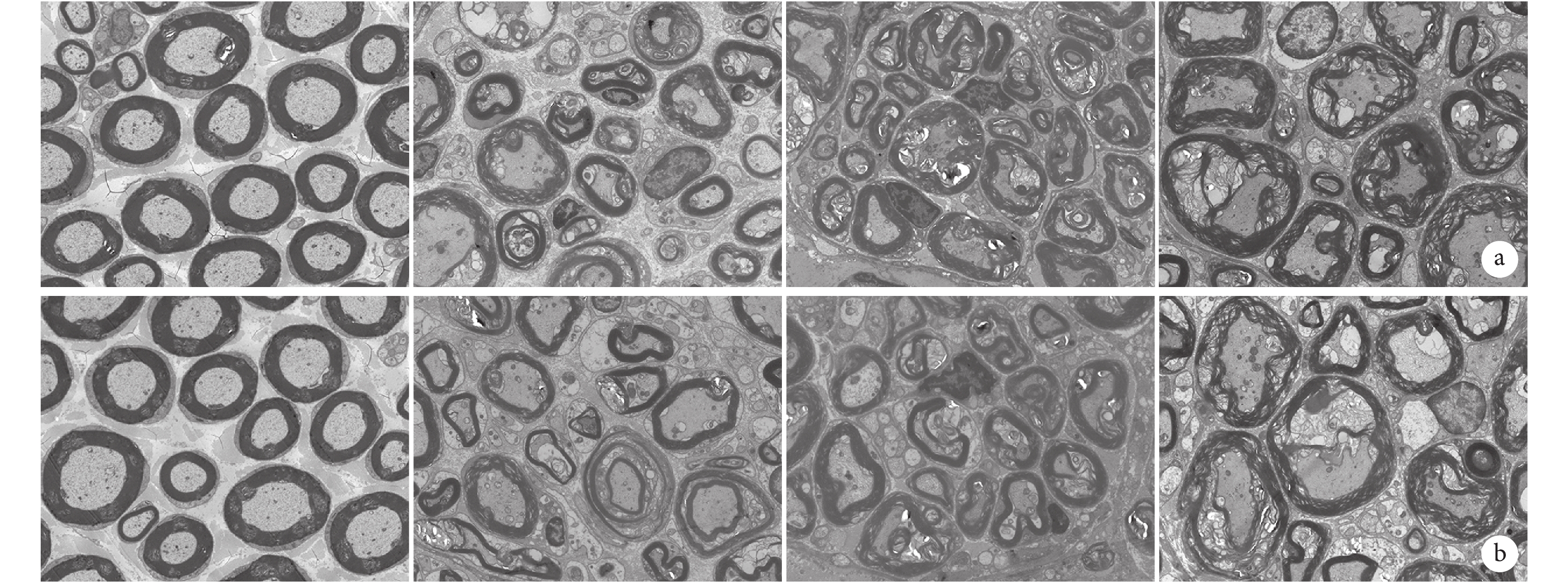 圖5
透射電鏡觀察術后各時間點坐骨神經近、遠側斷端超微結構(×2 000)
圖5
透射電鏡觀察術后各時間點坐骨神經近、遠側斷端超微結構(×2 000)
從左至右分別為術后 0、1、4、7 d a. 近側斷端;b. 遠側斷端
Figure5. Ultrastructure of the proximal and distal stumps at different time points after operation observed by transmission electron microscopy (×2 000)From left to right for postoperative views at 0, 1, 4, and 7 days, respectively a. Proximal stump; b. Distal stump
3 討論
神經損傷,特別是中樞神經損傷難以再生,但 PNI 后神經元固有的再生能力被激活,促進再生微環境的形成,從而使神經斷端軸突及髓鞘再生,這一現象主要與雪旺細胞等的作用有關[5, 16]。雪旺細胞作為周圍神經特有的且最重要的膠質細胞,在神經損傷早期激化并大量增殖,參與近、遠側斷端一系列特異性改變,對 PNI 后修復過程具有決定性影響[17-20]。目前研究發現,PNI 后內源基因 PTEN 的活性受到 PI3K/Akt/mTOR 信號轉導途徑的負調控,并參與調節損傷產生的炎癥與免疫反應,影響雪旺細胞的遷移、增殖和分化,進而促進軸突、髓鞘再生等過程[21-22]。隨著基因芯片技術用于 PNI 的治療,miRNA 在調節真核基因表達中的作用變得尤為突出,這種平均 20~24 個核苷酸長度的非編碼單鏈 RNA 主要通過與靶基因 3′UTR 以完全或不完全互補結合方式發揮作用,阻遏翻譯過程或裂解靶 mRNA,抑制基因表達,實現對基因轉錄后表達的調控[23]。
大量基因芯片研究表明,PNI 后的修復過程中,絕大多數上調或下調基因的表達會在 1 周和 3、4 周出現 2 個極值[24]。本研究通過 PNI 后修復1 周內的 4 個不同時間點,對大鼠坐骨神經損傷后 miR-221 與 PTEN 之間是否存在相關性進行了探討。通過實時熒光定量 PCR 和 Western blot 分別檢測了近、遠側斷端 miR-221 以及 PTEN 的相對表達量,發現隨著神經損傷后時間增加,在神經斷端中 miR-221 呈上調趨勢,而 PTEN 呈顯著下調趨勢。為探究這一現象中二者關系,我們進行了雙熒光素酶報告基因實驗,結果顯示 PTEN 為 miR-221 的作用靶點,進而闡明了二者在 PNI 中存在負相關,即 PNI 后內源基因 PTEN 降低與 miR-221 上調有密不可分的聯系。
在此基礎上,本研究進一步分析了 PNI 后二者在神經近、遠側斷端中的表達量是否具有差異性。實時熒光定量 PCR、Western blot 檢測結果示 miR-221 及 PTEN 相對表達量在神經近、遠側斷端間存在著不同程度差異,隨著神經損傷時間延長,二者在近側斷端變化幅度大于遠側斷端,呈現的差異性趨勢也逐漸增大。對于這一現象,我們從形態學進行了觀察。通過免疫熒光染色在熒光顯微鏡下觀察 PTEN 蛋白在神經斷端橫截面上的定位分布及其陽性表達,提示神經近、遠側斷端存在的差異性。結合透射電鏡形態學超微結構觀察,從術后 4 d 開始,神經近側斷端中雪旺細胞的分裂、增殖程度明顯大于遠側斷端,且軸突、髓鞘的破壞程度小于遠側斷端,至 7 d 時達峰值。這種損傷早期發生的軸突、髓鞘崩解,雪旺細胞增殖以及產生的近、遠端修復差異性,闡述了 PNI 后的修復過程。對于神經近、遠側斷端 miR-221 和 PTEN 表達差異的產生,我們認為可能與神經的不對稱修復相關。遠側斷端的軸突由于得不到胞體的營養支持,僅能存活數天,之后很快發生變性、解體;近側斷端的軸突和髓鞘雖可有同樣變化,但潰變一般只到鄰近斷端的第 1 個側支處中止,僅出現 1 個或幾個郎飛結的變性[25]。然而,神經元胞體是否存在對近側斷端的營養支持及其某些特定的信號調節等作用,使得 miR-221 與 PTEN 在神經近、遠側斷端含量上存在差異性,均有待后續研究解決。
綜上述,本研究結果顯示 PNI 后的修復過程中 miR-221 表達上調靶向調控 PTEN 表達降低,從而產生近、遠側斷端 miR-221 及 PTEN 表達差異,這一現象可能參與 PNI 后的一系列修復過程。但PNI 后的修復是一個多因素共同參與的復雜過程,具體機制有待進一步研究。
作者貢獻:張臣鳴負責本研究具體實施、文章撰寫等所有工作;李宇、張玉強參與實驗模型構建、標本取材、實驗檢測、數據整理;曹宇、龔超、王晨亮參與動物飼養與管理、標本取材以及數據收集;王偉參與課題設計、組織以及指導。
利益沖突:作者聲明,本課題研究和文章撰寫過程中不存在任何利益沖突。基金項目經費支持沒有影響文章觀點和對研究數據客觀結果的統計分析及其報道。
機構倫理問題:本研究通過錦州醫科大學動物倫理學委員會批準。所有實驗大鼠符合國家一級動物標準,實驗動物生產許可證號:SCXK-遼 2017-0003,使用許可證號:SYXK-遼 2014-0002。
周圍神經損傷(peripheral nerve injury,PNI)后經典的修復過程是神經遠側斷端瓦勒變性溶解消失,重新由近側斷端向遠處生長出整個遠側神經[1-4]。大量研究證明,磷酸酶與張力蛋白同源物(phosphatase and tension protein homologue,PTEN)在 PNI 后的修復中起重要作用,其蛋白表達量降低可以促進軸突及髓鞘再生[5-7]。近年來,微小 RNA(microRNA,miRNA)因在各種生物學功能中的調節作用引起了極大關注。有研究證實,在多種腫瘤細胞中,miR-221 對 PTEN 具有調控作用[8-10],其通過與 PTEN mRNA 3′端非翻譯區(3′UTR)互補結合來抑制 PTEN 磷酸水解酶表達,從而減弱 PTEN 在控制細胞增殖、凋亡、轉移與侵襲中的效能[11]。然而,miR-221/PTEN 在 PNI 后修復中是否存在相關性未見報道。此外,PNI 后修復受到神經遠側斷端發出的信號調控[12-13],這種不對稱修復現象使軸突再生由神經近側斷端向遠側斷端生長[14],miR-221/PTEN 在近、遠側斷端表達變化是否存在差異尚未明確。本實驗中,我們旨在研究 miR-221 與 PTEN 在 PNI 后修復中的表達及調控關系,比較近、遠側斷端中二者表達差異,從而為臨床診治 PNI 提供新靶點。
1 材料與方法
1.1 實驗動物、細胞及主要試劑、儀器
SPF 級雄性 SD 大鼠 96 只,體質量(200±20)g,由錦州醫科大學實驗動物中心提供,于光照和黑暗每 12 小時交替環境中標準飼養。293T 細胞(ATCC 細胞庫,美國)。
FBS(Serapro 公司,美國);DMEM 培養基、青/鏈霉素、胰蛋白酶(HyClone,美國);Opti-MEM 培養基 (GIBCO 公司,美國);兔抗鼠 PTEN 多克隆抗體(北京博奧森生物技術有限公司);兔抗鼠 GAPDH 多克隆抗體、辣根過氧化物酶標記的山羊抗兔 IgG 二抗(Earthox 公司,美國);Alexa Fluor 647-結合羊抗兔二抗(Abcam 公司,英國);牛血清白蛋白(Sigma 公司,美國);蛋白提取試劑盒、BCA 蛋白定量試劑盒、SDS-PAGE 凝膠試劑盒、ECL 化學發光試劑盒(上海碧云天生物技術有限公司);蛋白 Marker、GeneJETTMRNA 純化試劑盒(Thermo Fisher 公司,美國);GoScriptTM Reverse Transcription System 逆轉錄試劑盒、GoTaq? qPCR Master Mix 試劑盒、Dual-Luciferase? Reporter Assay System 試劑盒、pmirGLO Dual-Luciferase? miRNA Target Expression Vector(Promega 公司,美國);miR-221 模擬物(蘇州吉瑪基因股份有限公司);LipofectamineTM 2000(Invitrogen 公司,美國);QuikChange? Lightning Site-Directed Mutagenesis Kits 試劑盒(Stratagene 公司,美國);Gluta 固定液(Solarbio 公司,美國)。
顯微外科手術器械(泗洪金睛醫療器械有限公司);病理切片機、超薄切片機(Leica 公司,德國);正置熒光顯微鏡(Nikon 公司,日本);紫外分光光度計(Tecan 公司,美國);全自動電泳儀、化學發光凝膠成像系統(Bio-Rad 公司,美國);ABI stepone plus 實時熒光定量 PCR 儀(Applied Biosystems 公司,美國);透射電鏡(Hitachi 公司,日本)。
1.2 細胞培養
取 293T 細胞采用含 10%FBS 的 DMEM 培養基培養,置于 37 ℃、5%CO2 細胞孵育箱中培養。
1.3 大鼠坐骨神經損傷模型建立
取 96 只 SD 大鼠環境適應處理 12 h 后,100 g/L 水合氯醛(350 mg/kg)腹腔注射麻醉;麻醉后靜置動物約 3 min,備皮后于右下肢股骨中段下緣 5 mm 處作平行于股骨的切口約 5 mm[15],組織鉗鈍性分離筋膜及肌間隙后,暴露坐骨神經,用顯微外科剪離斷坐骨神經,觀察動物神經反射情況。逐層縫合后注射阿米卡星 0.1 mL 預防感染,切口包扎。待麻醉復蘇后,觀察動物運動恢復情況。分別于術后 0(術后即刻)、1、4、7 d,以麻醉后頸椎脫臼法分別處死 24 只大鼠,無菌條件下取坐骨神經近、遠側斷端組織各 5 mm 進行以下觀測。
1.4 觀測指標
1.4.1 實時熒光定量PCR檢測miR-221基因表達
于術后各時間點分別取 6 只大鼠坐骨神經近、遠側斷端組織,液氮速凍后研磨成粉,用 RNA 純化試劑盒過柱法提取組織總 RNA,經紫外分光光度計檢測,測定吸光度(A)值(A260/A280 值在 1.9~2.2 之間用于 cDNA 合成)。按照逆轉錄反應體系進行逆轉錄,所得 cDNA 在實時熒光定量反應體系下擴增,擴增 40 個循環。引物序列:miR-221 上游 5′-ATGCTCTAAGCTACATTGTCTGC-3′,下游 5′-TATGGTTGTTCTGCTCTCTGTCTC-3′;內參照 U6 上游 5′-CGCTTCGGCAGCACATATAC-3′,下游 5′-TTCACGAATTTGCGTGTCATC-3′。采用 2?ΔΔCt法計算 miR-221 相對表達量,實驗重復 3 次,取均值。
1.4.2 Western blot檢測PTEN蛋白表達
于術后各時間點分別取 6 只大鼠坐骨神經近、遠側斷端組織,于冰上按 1 mg 神經組織∶20 μL RIPA 裂解液進行裂解,4 ℃ 冰浴勻漿 30 min 后,以離心半徑 3 cm、12 000 r/min 離心 30 min,取上清液即為所需蛋白液。BCA 法蛋白定量后調整至同等濃度,每孔上樣 30 μg,經 10%SDS-PAGE 凝膠電泳后轉移至聚偏二氟乙烯膜,1% 牛血清白蛋白封閉 1 h;加入 TBST 稀釋兔抗鼠 PTEN 多克隆抗體(1︰1 000)、兔抗鼠 GAPDH 多克隆抗體(1︰3 000),4 ℃ 孵育過夜;TBST 洗膜 3 次,加入辣根過氧化物酶標記的羊抗兔 IgG 二抗(1︰10 000)室溫孵育 2 h;TBST 洗膜 3 次,在暗室內行化學發光劑 ECL 反應,化學發光凝膠成像系統取像并測定條帶灰度值。以 PTEN 蛋白條帶和 GAPDH 內參條帶灰度值的比值作為目的蛋白的相對表達量。
1.4.3 雙熒光素酶報告基因實驗
通過 Targetscan、miRDB、Miranda 和 PicTar 等生物信息學軟件預測 miR-221 靶向結合 PTEN 3′UTR 的位點進行分析。利用 PCR 擴增包含 miR-221 結合位點的 PTEN 3′UTR 序列,將其克隆到 pmirGLO 報告載體,構建野生型 pmirGLO-PTEN-WT 報告載體;用單點突變試劑盒突變 pmirGLO-PTEN-WT 上的 miR-221 結合位點,構建突變型 pmirGLO-PTEN-Mut 報告載體。取對數生長期 293T 細胞,以 1.5×104個/孔接種于 96 孔板中。將 Opti-MEM 培養基稀釋的野生型和突變型報告載體分別與 miR-221 模擬物(miR-221 mimic)或 miR-陰性對照(miR-NC)混合,混合后加入 LipofectamineTM 2000 試劑。將所得混合液分別與 Opti-MEM 培養基以 1∶1 比例加入 293T 細胞,其轉染濃度報告載體為每孔 250 ng,miR-221 mimic 及 NC 為每孔 50 nmol/L 孔。培養 6 h 后再加 100 μL Opti-MEM 培養基,48 h 后用 Dual-Luciferase? Reporter Assay System 試劑盒檢測各組細胞螢火蟲熒光酶熒光值(M1)及海腎熒光素酶熒光值(M2),以 M1/M2 比值作為相對熒光值。
1.4.4 免疫熒光染色觀察PTEN蛋白表達
于術后各時間點分別取 6 只大鼠坐骨神經近、遠側斷端組織,于 4% 多聚甲醛固定 24 h,行脫水、石蠟包埋、切片等處理,常規行抗原修復、山羊血清封閉后,滴加兔抗鼠 PTEN 多克隆抗體(1∶400)4 ℃ 孵育過夜;置于搖床 PBS 洗滌 3 次,Alexa Fluor 647-結合羊抗兔二抗(1∶500)避光室溫孵育 30 min;DAPI 染液處理后,洗滌并用抗熒光淬滅封片劑封片,正置熒光顯微鏡下觀察 PTEN 蛋白在神經斷端橫截面上的分布。取 100 倍鏡下單個神經橫截面視野,應用 Image-Pro Plus 6.0 軟件分析其平均吸光度(IA)值作為 PTEN 蛋白相對表達量。
1.4.5 透射電鏡觀察
于術后各時間點分別取 6 只大鼠坐骨神經近、遠側斷端組織,取 1 mm×1 mm×1 mm 神經組織,Gluta 固定液固定 1~4 h,再用 1% 鋨酸-0.1 mol/L PBS 室溫固定 2 h,脫水、滲透、包埋后制作厚 60 nm 超薄切片,經鈾鉛雙染色后透射電鏡觀察神經軸突、髓鞘等超微結構的形態學變化。
1.5 統計學方法
采用 SPSS19.0 統計軟件進行分析。數據以均數±標準差表示,同側斷端各時間點間比較采用單因素方差分析,兩兩比較采用 Bonferroni 法;左、右側斷端同一時間點間比較采用獨立樣本 t 檢驗;雙熒光素酶報告基因實驗組間比較采用 Dunnett’s t 檢驗;檢驗水準 α=0.05。
2 結果
2.1 實時熒光定量PCR檢測miR-221表達
隨時間延長,大鼠坐骨神經近、遠側斷端組織中 miR-221 相對表達量均逐漸增加,術后各時間點間比較差異均有統計學意義(P<0.05)。術后 1、4、7 d 近側斷端組織中 miR-221 相對表達量均顯著高于遠側斷端,差異有統計學意義(P<0.05)。見圖1。
圖1
實時熒光定量PCR檢測術后各時間點大鼠坐骨神經 近、遠側斷端組織中miR-221相對表達量
Figure1.
The relative expression of miR-221 in the proximal and distal stumps at different time points after operation by real-time fluorescent quantitative PCR
2.2 Western blot檢測PTEN蛋白表達
隨時間延長,大鼠坐骨神經近、遠側斷端組織中 PTEN 蛋白相對表達量均逐漸減少,術后各時間點間比較差異均有統計學意義(P<0.05)。術后 1、4、7 d 近側斷端組織中 PTEN 蛋白相對表達量均顯著低于遠側斷端,差異有統計學意義(P<0.05)。見圖2。
圖2
Western blot檢測術后各時間點大鼠坐骨神經近、遠側斷端組織中PTEN蛋白表達
Mr:相對分子質量 1:0 d 2:1 d 3:4 d 4:7 d a. 近側斷端蛋白條帶圖;b. 遠側斷端蛋白條帶圖;c. PTEN 蛋白相對表達量
Figure2. The expression of PTEN protein in the proximal and distal stumps at different time points after operation detected by Western blotMr: Relative molecular mass 1: 0 day 2: 1 day 3: 4 days 4: 7 days; a. Protein bands of the proximal stump; b. Protein bands of the distal stump; c. Relative expression of PTEN protein
2.3 雙熒光素酶報告基因實驗
生物信息預測結果表明,miR-221 序列上存在 PTEN 的連續結合序列,見表1。通過構建野生型和突變型報告載體,雙熒光素酶報告基因實驗發現,與 miR-NC(1.003±0.009)相比,miR-221 mimic 與野生型報告載體共轉染時顯著降低了熒光素酶相對活性(0.402±0.045),差異有統計學意義(t=15.78,P=0.00);而與突變型報告載體共轉染時恢復了熒光素酶相對活性(0.972±0.021),差異無統計學意義(t=1.86,P=0.14)。
2.4 免疫熒光染色觀察PTEN蛋白表達
熒光顯微鏡觀察示,PTEN 蛋白主要表達于細胞質,部分細胞核中也有少許表達。隨神經損傷后修復時間的延長,PTEN 陽性表達明顯減少,其亞細胞定位無明顯變化。見圖3。
圖3
免疫熒光染色觀察術后各時間點坐骨神經近、遠側斷端組織中PTEN蛋白表達(熒光顯微鏡×100)
從左至右依次為術后 0、1、4、7 d a. 近側斷端;b. 遠側斷端
Figure3. Immunofluorescence staining observation of the expression of PTEN protein in the proximal and distal stumps at different time points after operation (Fluorescence microscopy×100)From left to right for postoperative views at 0, 1, 4, and 7 days, respectively a. Proximal stump; b. Distal stump
定量檢測示隨時間延長,大鼠坐骨神經近、遠側斷端組織 IA 值均逐漸減小,術后各時間點間比較差異均有統計學意義(P<0.05)。術后 1、4、7 d 遠側斷端組織 IA 值均顯著高于近側斷端,差異有統計學意義(P<0.05)。見圖4。
圖4
免疫熒光染色檢測術后各時間點坐骨神經近、遠側斷端 組織IA值
Figure4.
Detection of the IA value in the proximal and distal stumps at different time points after operation by immuno fluorescence staining
2.5 透射電鏡觀察
術后 0 d,近、遠側斷端正常軸突、髓鞘等超微結構形態規整、結構清晰,無病理改變。術后 1 d,近、遠側斷端雪旺細胞增殖及潰變均不明顯,髓鞘整體形態結構與術后 0 d 接近,但髓鞘板層略有疏松,鞘內軸漿顆粒消失。術后 4 d,雪旺細胞較 0 d 時明顯增殖旺盛,呈較明顯神經潰變,近、遠側斷端可見大量微管及微絲崩解,線粒體腫脹,髓鞘開始破壞,髓鞘內板層結構模糊,間隙增寬、松散,形態不規則;與遠側斷端相比,近側斷端雪旺細胞分裂、增殖更明顯,且軸突、髓鞘破壞程度更小。術后 7 d,已無完整髓鞘結構,雪旺細胞遷移引導軸突再生,近、遠側斷端神經潰變達到高峰,且遠側斷端神經潰變較近側斷端更徹底;近側斷端可見少數再生軸突已被雪旺細胞包繞,以及少數新生神經纖維及未成熟的新生髓鞘。見圖5。
圖5
透射電鏡觀察術后各時間點坐骨神經近、遠側斷端超微結構(×2 000)
從左至右分別為術后 0、1、4、7 d a. 近側斷端;b. 遠側斷端
Figure5. Ultrastructure of the proximal and distal stumps at different time points after operation observed by transmission electron microscopy (×2 000)From left to right for postoperative views at 0, 1, 4, and 7 days, respectively a. Proximal stump; b. Distal stump
3 討論
神經損傷,特別是中樞神經損傷難以再生,但 PNI 后神經元固有的再生能力被激活,促進再生微環境的形成,從而使神經斷端軸突及髓鞘再生,這一現象主要與雪旺細胞等的作用有關[5, 16]。雪旺細胞作為周圍神經特有的且最重要的膠質細胞,在神經損傷早期激化并大量增殖,參與近、遠側斷端一系列特異性改變,對 PNI 后修復過程具有決定性影響[17-20]。目前研究發現,PNI 后內源基因 PTEN 的活性受到 PI3K/Akt/mTOR 信號轉導途徑的負調控,并參與調節損傷產生的炎癥與免疫反應,影響雪旺細胞的遷移、增殖和分化,進而促進軸突、髓鞘再生等過程[21-22]。隨著基因芯片技術用于 PNI 的治療,miRNA 在調節真核基因表達中的作用變得尤為突出,這種平均 20~24 個核苷酸長度的非編碼單鏈 RNA 主要通過與靶基因 3′UTR 以完全或不完全互補結合方式發揮作用,阻遏翻譯過程或裂解靶 mRNA,抑制基因表達,實現對基因轉錄后表達的調控[23]。
大量基因芯片研究表明,PNI 后的修復過程中,絕大多數上調或下調基因的表達會在 1 周和 3、4 周出現 2 個極值[24]。本研究通過 PNI 后修復1 周內的 4 個不同時間點,對大鼠坐骨神經損傷后 miR-221 與 PTEN 之間是否存在相關性進行了探討。通過實時熒光定量 PCR 和 Western blot 分別檢測了近、遠側斷端 miR-221 以及 PTEN 的相對表達量,發現隨著神經損傷后時間增加,在神經斷端中 miR-221 呈上調趨勢,而 PTEN 呈顯著下調趨勢。為探究這一現象中二者關系,我們進行了雙熒光素酶報告基因實驗,結果顯示 PTEN 為 miR-221 的作用靶點,進而闡明了二者在 PNI 中存在負相關,即 PNI 后內源基因 PTEN 降低與 miR-221 上調有密不可分的聯系。
在此基礎上,本研究進一步分析了 PNI 后二者在神經近、遠側斷端中的表達量是否具有差異性。實時熒光定量 PCR、Western blot 檢測結果示 miR-221 及 PTEN 相對表達量在神經近、遠側斷端間存在著不同程度差異,隨著神經損傷時間延長,二者在近側斷端變化幅度大于遠側斷端,呈現的差異性趨勢也逐漸增大。對于這一現象,我們從形態學進行了觀察。通過免疫熒光染色在熒光顯微鏡下觀察 PTEN 蛋白在神經斷端橫截面上的定位分布及其陽性表達,提示神經近、遠側斷端存在的差異性。結合透射電鏡形態學超微結構觀察,從術后 4 d 開始,神經近側斷端中雪旺細胞的分裂、增殖程度明顯大于遠側斷端,且軸突、髓鞘的破壞程度小于遠側斷端,至 7 d 時達峰值。這種損傷早期發生的軸突、髓鞘崩解,雪旺細胞增殖以及產生的近、遠端修復差異性,闡述了 PNI 后的修復過程。對于神經近、遠側斷端 miR-221 和 PTEN 表達差異的產生,我們認為可能與神經的不對稱修復相關。遠側斷端的軸突由于得不到胞體的營養支持,僅能存活數天,之后很快發生變性、解體;近側斷端的軸突和髓鞘雖可有同樣變化,但潰變一般只到鄰近斷端的第 1 個側支處中止,僅出現 1 個或幾個郎飛結的變性[25]。然而,神經元胞體是否存在對近側斷端的營養支持及其某些特定的信號調節等作用,使得 miR-221 與 PTEN 在神經近、遠側斷端含量上存在差異性,均有待后續研究解決。
綜上述,本研究結果顯示 PNI 后的修復過程中 miR-221 表達上調靶向調控 PTEN 表達降低,從而產生近、遠側斷端 miR-221 及 PTEN 表達差異,這一現象可能參與 PNI 后的一系列修復過程。但PNI 后的修復是一個多因素共同參與的復雜過程,具體機制有待進一步研究。
作者貢獻:張臣鳴負責本研究具體實施、文章撰寫等所有工作;李宇、張玉強參與實驗模型構建、標本取材、實驗檢測、數據整理;曹宇、龔超、王晨亮參與動物飼養與管理、標本取材以及數據收集;王偉參與課題設計、組織以及指導。
利益沖突:作者聲明,本課題研究和文章撰寫過程中不存在任何利益沖突。基金項目經費支持沒有影響文章觀點和對研究數據客觀結果的統計分析及其報道。
機構倫理問題:本研究通過錦州醫科大學動物倫理學委員會批準。所有實驗大鼠符合國家一級動物標準,實驗動物生產許可證號:SCXK-遼 2017-0003,使用許可證號:SYXK-遼 2014-0002。