引用本文: 席懿璇, 竇國睿, 周子義, 常天芳, 儲昭節. 嚴重急性呼吸綜合征冠狀病毒2型感染對小鼠視網膜光感受器細胞形態、增殖、凋亡及免疫功能的影響. 中華眼底病雜志, 2024, 40(10): 772-780. doi: 10.3760/cma.j.cn511434-20240409-00148 復制
版權信息: ?四川大學華西醫院華西期刊社《中華眼底病雜志》版權所有,未經授權不得轉載、改編
由嚴重急性呼吸綜合征冠狀病毒2型(SARS-CoV-2)引起的新型冠狀病毒感染(COVID-19)是一種嚴重的疾病[1-2]。SARS-COV-2感染后會促進炎癥反應,引起高凝狀態,導致多種全身并發癥,其入侵視網膜可引起視網膜炎癥及其一系列眼部并發癥,如結膜炎、視網膜出血、視網膜靜脈阻塞、急性黃斑神經視網膜病變(AMN)等[3-6]。AMN是一種多發生于年輕、健康女性的較為罕見的視網膜疾病,其主要癥狀為患者眼部驟然出現1個或多個旁中心暗點,視力正常或輕度下降,黃斑區損傷較大的患者,可導致視力暫時或永久性下降[7]。AMN誘發因素包括口服避孕藥、呼吸系統損傷、流感樣疾病和咖啡因攝入等[8-9]。SARS-CoV-2感染和疫苗接種對AMN發生和發展可能具有一定影響[10-12]。SARS-CoV-2通過尖峰(S)蛋白與其受體血管緊張素轉化酶2(ACE2)結合的方式侵入眼部[13-14]。AMN患者視網膜敏感度降低的區域內,光感受器結構優先破壞,提示視網膜光感受器損傷與AMN發病機制相關[15]。然而,目前關于SARS-CoV-2在視網膜光感受器的研究鮮見報道。本研究通過體外細胞實驗分析SARS-CoV-2對小鼠光感受器細胞(661w細胞)的形態、增殖、凋亡、周期及免疫功能的作用,并初步探討SARS-CoV-2引發AMN的潛在機制。現將結果報道如下。
1 材料和方法
1.1 材料
小鼠光感受器細胞株(661w細胞,上海通派生物科技有限公司);Dulbecco改良Eagle培養基(DMEM,L540KJ)、4',6-二脒基-2-苯基吲哚(DAPI,C0065)(美國Gibco公司);兔抗小鼠腫瘤壞死因子(TNF)-α多克隆抗體(ab307164)、兔抗小鼠B細胞淋巴瘤-2(Bcl-2)相關X蛋白(Bax)單克隆抗體(ab32503)(英國Abcam公司);兔抗小鼠白細胞介素(IL)-6多克隆抗體、小鼠抗小鼠Bcl-2單克隆抗體、兔抗小鼠β-肌動蛋白(actin)、山羊抗兔免疫球蛋白(Ig)G二抗、山羊抗小鼠IgG二抗(武漢三鷹生物技術有限公司);Alexa Fluor 594山羊抗兔IgG二抗、異硫氰酸熒光素(FITC)標記的鬼筆環肽(40735ES75)、細胞凋亡及周期試劑盒(40302E)(上海翌圣生物科技股份有限公司);兔抗小鼠ACE2單克隆抗體(PA5-47488,美國Thermo Fisher公司);Triton X-100(上海生工生物工程股份有限公司);ACE2 siRNA過表達慢病毒(siACE2)(上海吉凱公司);SARS-CoV-2假病毒(PSV005,北京依翹神州生物技術公司);細胞計數試劑盒8(CCK-8,美國Selleck公司);磷酸鹽緩沖液(PBS,北京蘭杰柯科技有限公司);DMEM高糖完全培養基、胰蛋白酶、20倍洗膜緩沖液(TBST)(北京索萊寶科技有限公司);酶聯免疫檢測儀(ELX800,美國Bio-Teck公司)。正置光學顯微鏡(日本Olympus公司);iQ5實時熒光定量聚合酶鏈反應(qPCR)儀、細胞流式檢測儀(美國Thermo Fisher公司);FV3000型激光掃描共聚焦顯微鏡(日本Nikon公司)。
1.2 方法
細胞培養和分組。661w細胞株置于含10%胎牛血清和1%雙抗(青霉素/鏈霉素)的DMEM培養基中,于37℃、含5% CO2的細胞培養箱中培養至貼壁,取對數生長期細胞用于實驗。將661w細胞分為三組,分別為正常組(未經任何處理)、siACE2組(過表達ACE2且未感染假病毒)及感染組(過表達ACE2并感染假病毒),其中感染組再分為5 TU/ml假病毒組、15 TU/ml假病毒組、30 TU/ml 假病毒組、50 TU/ml假病毒組,分別誘導12、24、48、72 h。
構建siACE2過表達光感受器細胞。應用DMEM高糖完全培養基培養661w細胞,在37℃且含5% CO2的細胞培養箱中培養至貼壁。將細胞以1×104個/ml的細胞密度接種于24孔板中,細胞貼壁后,加入1×108 TU/ml siACE2轉染8 h,更換完全培養基,繼續培養48~72 h,細胞密度達到70%~80%時,0.25%不含乙二胺四乙酸(EDTA)胰酶消化細胞,以離心半徑15cm,1 200 r/min離心5 min,完全培養基重懸細胞,以1×105個/ml的661w細胞密度接種于12孔板中,培養至貼壁后,更換含有3 μg/ml Puromycin的DMED完全培養基繼續培養48 h,將ACE2過表達的661w細胞進行擴增及凍存保種。
激光共聚焦顯微鏡檢測ACE2轉染效率。將siACE2轉染后的細胞以5×103個/ml的細胞密度接種于共聚焦細胞培養皿中培養;細胞培養至50%后,4%多聚甲醛固定30 min,PBS漂洗3次,10 min/次;0.5% Triton X-100中孵育30 min,2%牛血清白蛋白封閉,室溫靜置1 h。加入兔抗小鼠ACE2一抗(1∶500),4 ℃過夜孵育,PBS漂洗3次;加入羊抗小鼠IgG抗體(1∶1 000),室溫孵育2 h,PBS漂洗3次,DAPI封片液封片,激光共聚焦顯微鏡下觀察拍照。
光學顯微鏡觀察各組細胞形態變化。將細胞以1×105個/ml的細胞密度接種于6孔板中,培養至細胞貼壁后,分別用假病毒誘導12、24、48、72 h,光學顯微鏡下觀察細胞形態并采集圖像。
CCK-8法檢測siACE組及感染組細胞活性。各組細胞以1×104個/ml的細胞密度接種于96孔板中,分別用假病毒誘導12、24、48、72 h,每孔滴加10 μl CCK-8試劑孵育1 h,酶鏈免疫檢測儀測量波長450 nm處吸光度[A,舊稱光密度(OD)]值,每組設3個復孔,實驗重復3次。計算不同病毒感染濃度及不同感染時間的細胞存活率,統計細胞增殖情況。細胞存活率=[A(感染)-A(空白)/A(對照)-A(空白)]×100%。
流式細胞儀檢測siACE組及感染組細胞凋亡情況。各組細胞以5×104個/ml的細胞密度接種于24孔板中,細胞貼壁后對各組細胞應用假病毒誘導12、24、48、72 h。0.25%無EDTA胰蛋白酶消化細胞,300 ×g離心5 min,加入膜聯蛋白(Annexin) V-FITC探針混勻,室溫避光孵育20min,加入碘化丙啶(PI)染色液混勻,冰浴避光孵育15 min。流式細胞儀檢測細胞凋亡情況,并使用 BDFACSuite軟件進行分析。凋亡指數=(早期凋亡細胞數量+晚期凋亡細胞數量)/總細胞數量。
流式細胞儀檢測siACE組、30 TU/ml假病毒組細胞周期。siACE組、30 TU/ml假病毒組細胞以1×105個/ml的細胞密度接種于24孔板中,假病毒誘導72 h,0.25%胰蛋白酶消化,加入預冷70%乙醇重懸細胞,4 ℃甲醛固定過夜,加入10 μl RNase A、10 μl PI,37°C下避光孵育30 min,流式細胞儀檢測細胞周期。
蛋白質免疫印跡法(Western blot)檢測siACE組、30 TU/ml假病毒組IL-6、TNF-α、Bcl-2、Bax蛋白相對表達量。siACE組、30 TU/ml假病毒組細胞以1×105個/ml的細胞密度接種于6孔板中,假病毒誘導72 h,提取各組細胞總蛋白,調整蛋白濃度。每個樣孔加入30~50 μg待測樣品,十二烷基硫酸鈉聚丙烯酰胺凝膠電泳中上樣電泳;80 V恒壓0.5 h,樣品進入分離膠后100 V恒壓2 h,300 mA轉膜2 h,5%脫脂牛奶常溫封閉2 h,一抗IL-6、TNF-α、Bax、Bcl-2、ACE2、β-actin(1∶1 000) 4℃過夜孵育(β-actin作為陽性對照),TBST洗膜10 min,重復3次;加入辣根過氧化物偶聯的二抗(1∶5 000),37℃孵育2 h,TBST洗膜10 min,重復5次;加入化學發光劑,暗室顯影成像。采用Image J Program軟件分析蛋白條帶灰度值,以目的條帶與β-actin 條帶的比值代表蛋白的相對表達水平。
qPCR檢測siACE組、30 TU/ml假病毒組細胞中IL-6、TNF-α、Bcl-2、Bax、核因子(NF)κB-1、NF-κB2以及NF- κB增強子(P65)、前體蛋白(P100)mRNA相對表達量。siACE組、30 TU/ml假病毒組細胞假病毒感染72 h,加入Trizol溶液裂解細胞;加入1/5總體積氯仿,充分混合并離心;將上層水相移至新管并加入等體積的異丙醇溶液,以離心半徑15 cm,12 000 r/min低溫離心10 min,移除上清液后加入無水乙醇再次低溫離心,倒出上清液,室溫放置干燥30 min,加入20 μl RNAse-free水,吹打溶解,提取各組細胞RNA,酶標儀檢測RNA濃度及純度;384孔板中依此加入2.5 μl cDNA、1 μl正向引物、1 μl反向引物和4 μl SYBR混合,加樣全程在冰上避光操作。以離心半徑7 cm、12 000 r/min離心10 min。引物序列由擎科生物公司設計(表1)。反應條件:50℃反應2 min,95℃變性10 min(1個循環);95℃變性15 s(40個循環);55℃擴增30 s;95℃反應15 s;60℃反應15 s,95℃ 反應15 s。置于qPCR儀中進行擴增并輸出循環閾值(Ct值),以β-actin為內參,依照公式2-ΔΔCt計算mRNA相對表達量。
采用GraphPad Prism v9.1.0.221軟件進行統計學分析。計量數據以均數±標準差(x±s)表示。多組間比較采用單因素方差分析;兩組間比較采用t檢驗。P<0.05為差異有統計學意義
2 結果
2.1 siACE2成功轉染至661w細胞
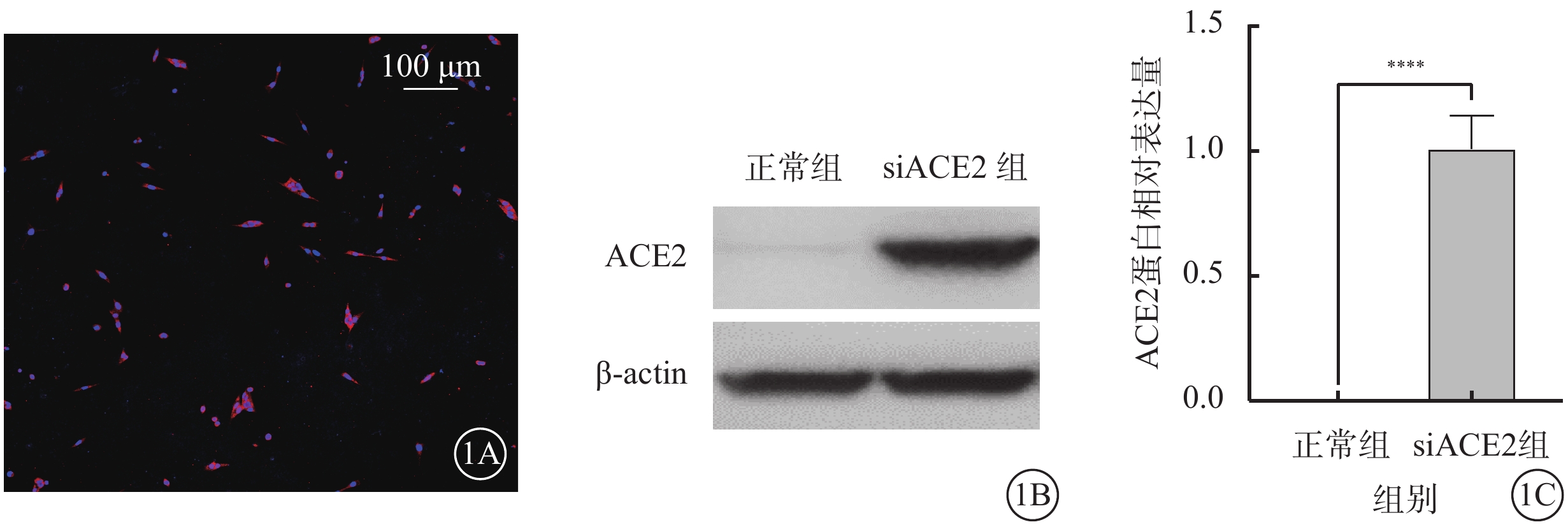
熒光顯微鏡觀察結果顯示,轉染siACE2 48 h后,661w細胞中ACE2轉染效率>90%(圖1A)。Western blot檢測結果顯示,與正常組比較,siACE組細胞中ACE2蛋白相對表達量顯著增加,差異有統計學意義(t=18.550,P<0.01)(圖1B,1C)。
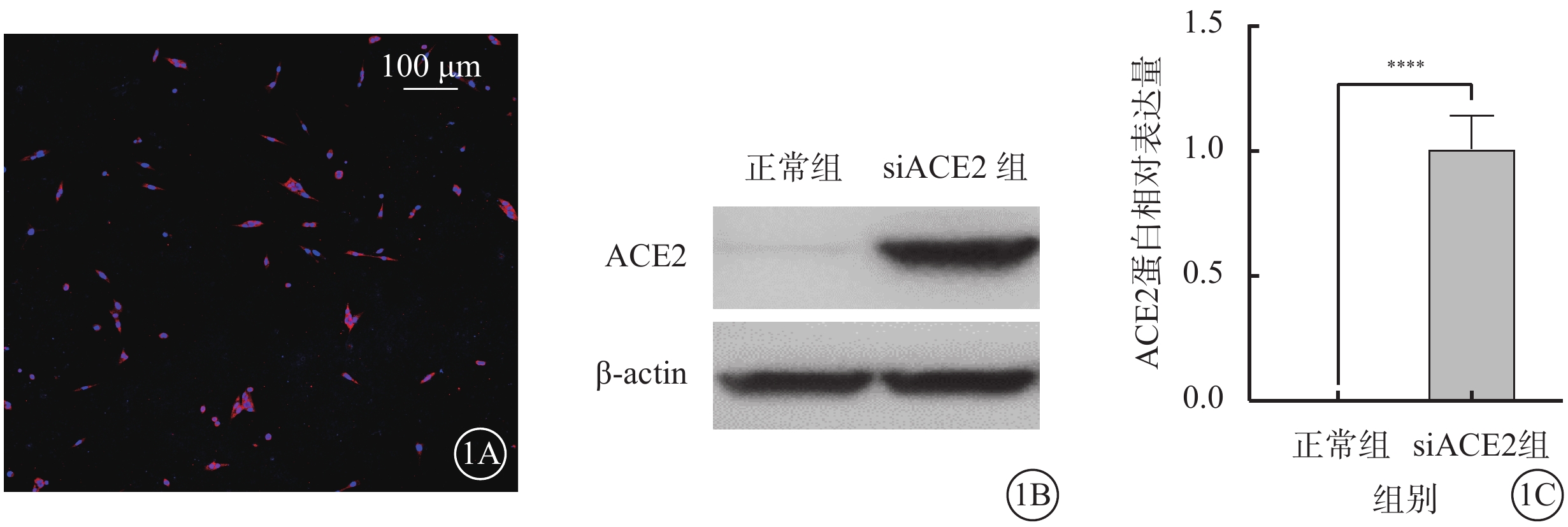 圖1
siACE2轉染效率驗證(n=3) 1A示661w細胞熒光顯微鏡像(DAPI,標尺:100 μm ),轉染siACE2 48 h后,ACE2轉染效率>90%;1B示電泳圖;1C示正常組、siACE組細胞中ACE2蛋白相對表達量比較, ****P<0.01 661w細胞:小鼠光感受器細胞;actin:肌動蛋白;ACE2:血管緊張素轉化酶2;siACE2:ACE2過表達慢病毒;DAPI:4',6-二脒基-2-苯基吲哚;正常組:661w細胞常規培養;siACE組:661w細胞轉染siACE2
圖1
siACE2轉染效率驗證(n=3) 1A示661w細胞熒光顯微鏡像(DAPI,標尺:100 μm ),轉染siACE2 48 h后,ACE2轉染效率>90%;1B示電泳圖;1C示正常組、siACE組細胞中ACE2蛋白相對表達量比較, ****P<0.01 661w細胞:小鼠光感受器細胞;actin:肌動蛋白;ACE2:血管緊張素轉化酶2;siACE2:ACE2過表達慢病毒;DAPI:4',6-二脒基-2-苯基吲哚;正常組:661w細胞常規培養;siACE組:661w細胞轉染siACE2
2.2 siACE組及感染組假病毒不同感染時間細胞形態
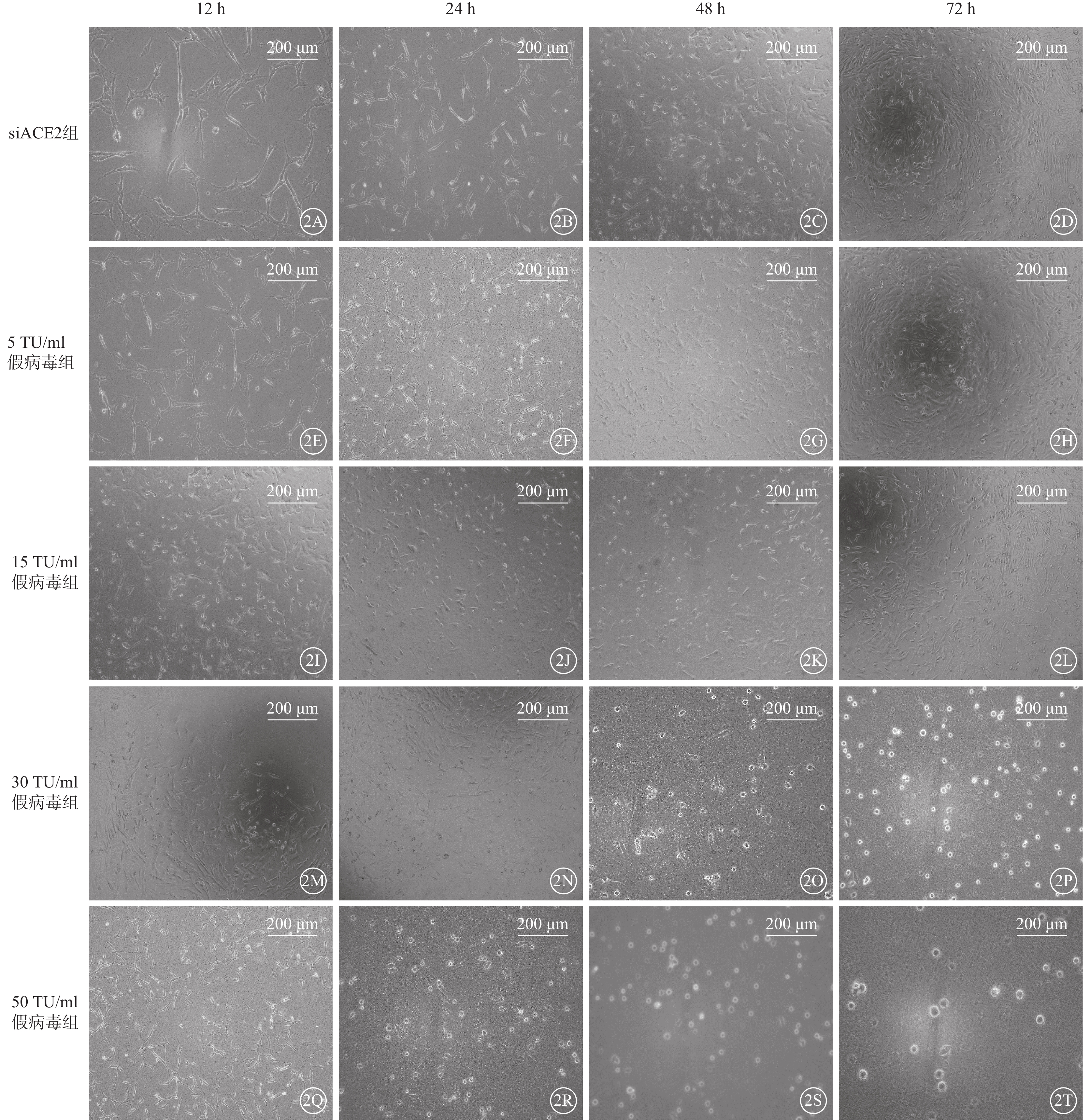
siACE2組細胞呈細長纖維狀,細胞形態清晰、成簇或平行排列(圖2A~2D);感染組細胞均出現不同程度皺縮(圖2E~2T)。與siACE組比較,5 TU/ml 假病毒組、15 TU/ml假病毒組細胞隨假病毒感染時間增加,細胞形態無明顯改變(圖2E~2L);72 h時,15 TU/ml 假病毒組細胞數量無明顯改變(圖2I~2L),部分細胞形態趨于圓形(圖2L)。30 TU/ml 假病毒組細胞形態隨假病毒誘導時間延長發生明顯改變;12 h、24 h時細胞形態無明顯改變,48 h時部分細胞形態出現皺縮,72 h時細胞嚴重皺縮,形態趨于圓形(圖2M~2P)。50 TU/ml 假病毒組細胞在假病毒誘導24 h時出現皺縮,隨時間延長逐漸嚴重,形態趨于圓形;72 h時細胞數量減少(圖2Q~2T)。
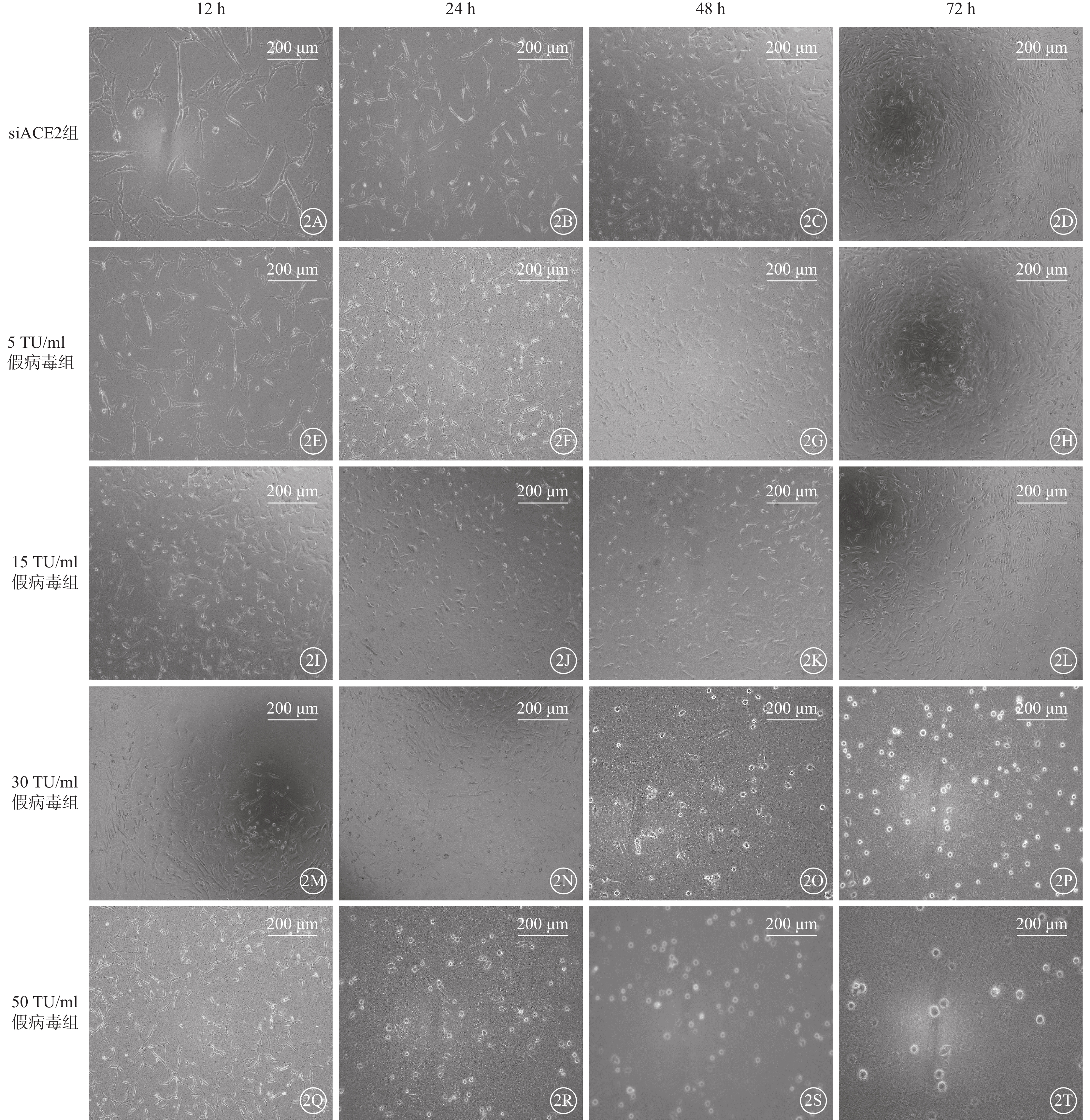 圖2
SARS-CoV-2假病毒不同誘導時間siACE2組與感染組細胞正置光學顯微鏡像(標尺:200 μm) 2A~2D、2E~2H、2I~2L、2M~2P、2Q~2T分別示siACE2組、5 TU/ml假病毒組、15 TU/ml假病毒組、30 TU/ml假病毒組、50 TU/ml假病毒組細胞假病毒誘導12 、24、48、72 h。siACE2組細胞呈細長纖維狀,細胞形態清晰、成簇或平行排列;感染組細胞均出現不同程度皺縮,隨假病毒轉染濃度、時間增加,細胞皺縮加劇,數量減少 661w細胞:小鼠光感受器細胞;SARS-CoV-2:急性呼吸綜合征冠狀病毒2型;ACE2:血管緊張素轉化酶2;siACE2:ACE2 siRNA過表達慢病毒;siACE2組:661w細胞轉染siACE2;感染組:661w細胞轉染siACE2后,分別加入5、15、30、50 TU/ml SARS-CoV-2假病毒誘導
圖2
SARS-CoV-2假病毒不同誘導時間siACE2組與感染組細胞正置光學顯微鏡像(標尺:200 μm) 2A~2D、2E~2H、2I~2L、2M~2P、2Q~2T分別示siACE2組、5 TU/ml假病毒組、15 TU/ml假病毒組、30 TU/ml假病毒組、50 TU/ml假病毒組細胞假病毒誘導12 、24、48、72 h。siACE2組細胞呈細長纖維狀,細胞形態清晰、成簇或平行排列;感染組細胞均出現不同程度皺縮,隨假病毒轉染濃度、時間增加,細胞皺縮加劇,數量減少 661w細胞:小鼠光感受器細胞;SARS-CoV-2:急性呼吸綜合征冠狀病毒2型;ACE2:血管緊張素轉化酶2;siACE2:ACE2 siRNA過表達慢病毒;siACE2組:661w細胞轉染siACE2;感染組:661w細胞轉染siACE2后,分別加入5、15、30、50 TU/ml SARS-CoV-2假病毒誘導
2.3 661w細胞存活率隨假病毒濃度增加、誘導時間延長而降低
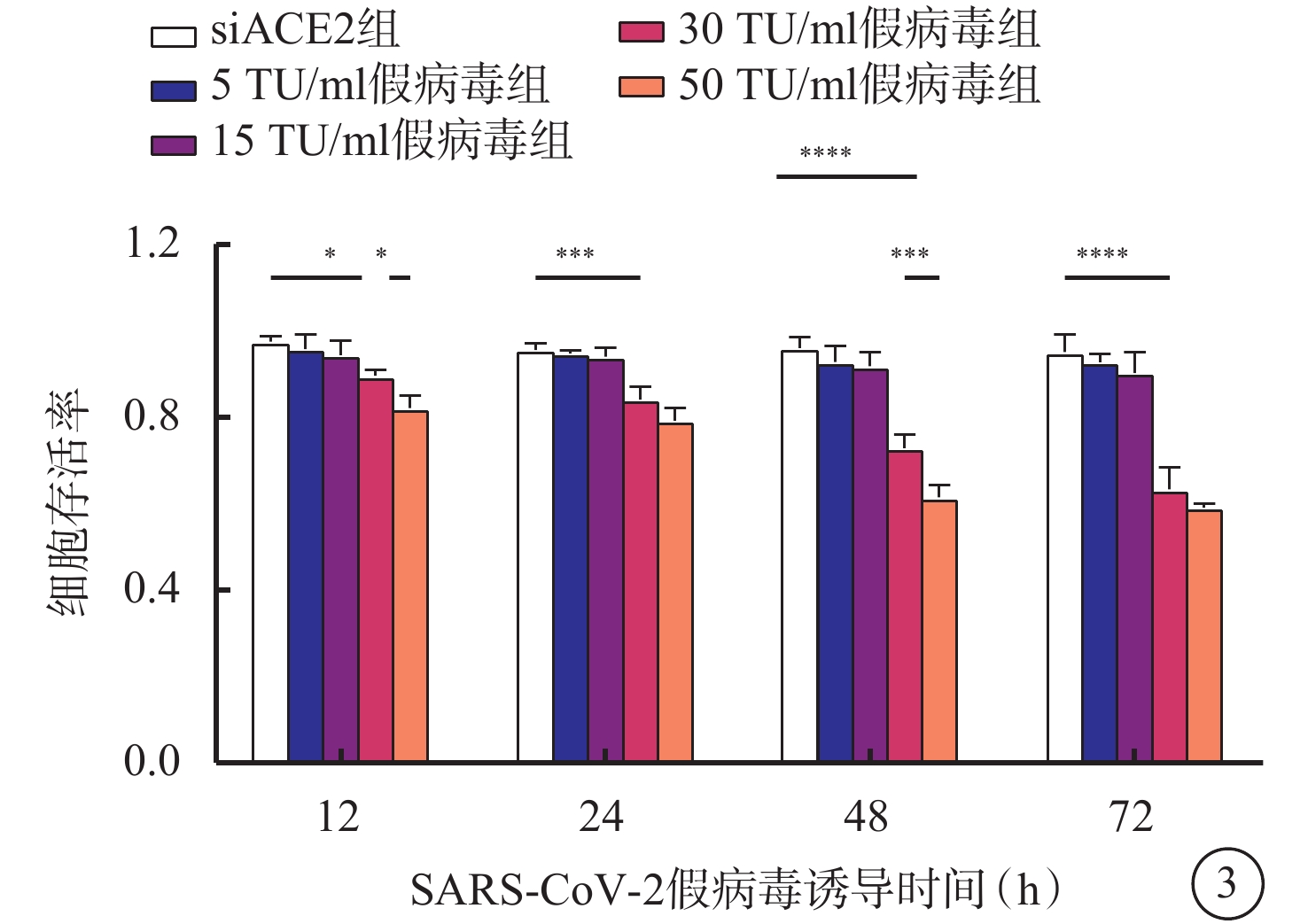
與siACE2組比較,感染組低濃度組(5 TU/ml假病毒組、15 TU/ml假病毒組)細胞誘導12、24、48、72 h后細胞存活率出現下調,但差異無統計學意義(F=0.840、0.412、1.498、1.138,P>0.05),感染組高濃度組(30 TU/ml假病毒組、50 TU/ml假病毒組)誘導12、24、48、72 h較siACE2組細胞存活率均降低,差異有統計學意義(F=3.304、4.772、9.629、13.220,P<0.05、0.01、0.01、0.01),而30 TU/ml假病毒組與50 TU/ml假病毒組誘導12、24、72 h比較,無顯著差異(t=3.113、2.030、1.787,P>0.05),但誘導48 h后,50 TU/ml假病毒組較30 TU/ml假病毒組細胞存活率明顯降低,差異有統計學意義(t=4.733,P<0.01)(圖3)。
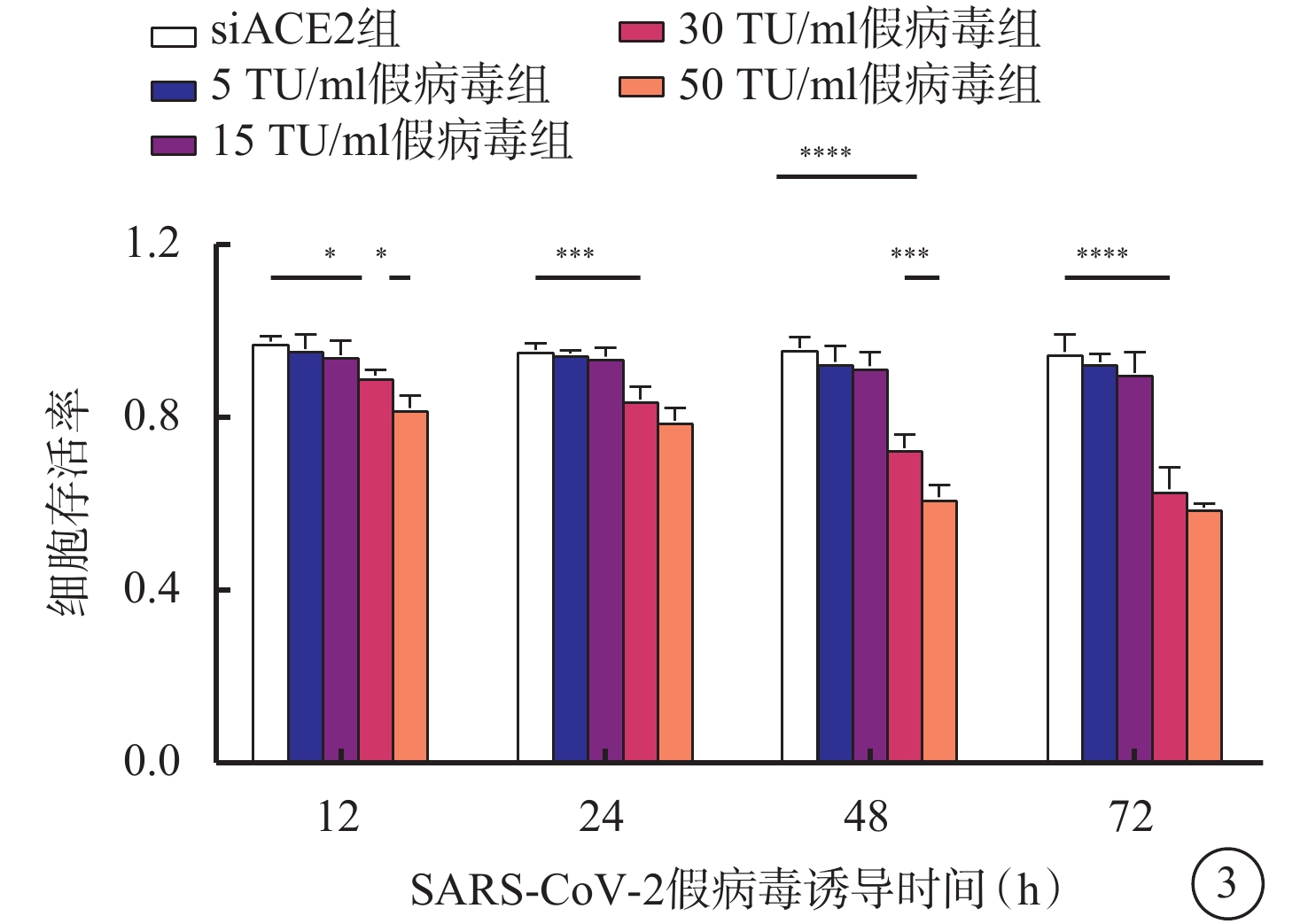 圖3
SARS-CoV-2假病毒不同誘導時間siACE2組與感染組細胞存活率比較(n=3) *P<0.05,** P<0.01 661w細胞:小鼠光感受器細胞;SARS-CoV-2:急性呼吸綜合征冠狀病毒2型;ACE2:血管緊張素轉化酶2;siACE2:ACE2 siRNA過表達慢病毒;siACE2組:661w細胞轉染siACE2;感染組:661w細胞轉染siACE2后,再分別加入5、15、30、50 TU/ml SARS-CoV-2假病毒誘導
圖3
SARS-CoV-2假病毒不同誘導時間siACE2組與感染組細胞存活率比較(n=3) *P<0.05,** P<0.01 661w細胞:小鼠光感受器細胞;SARS-CoV-2:急性呼吸綜合征冠狀病毒2型;ACE2:血管緊張素轉化酶2;siACE2:ACE2 siRNA過表達慢病毒;siACE2組:661w細胞轉染siACE2;感染組:661w細胞轉染siACE2后,再分別加入5、15、30、50 TU/ml SARS-CoV-2假病毒誘導
2.4 661w細胞凋亡指數隨SARS-CoV-2假病毒濃度、誘導時間增加而升高
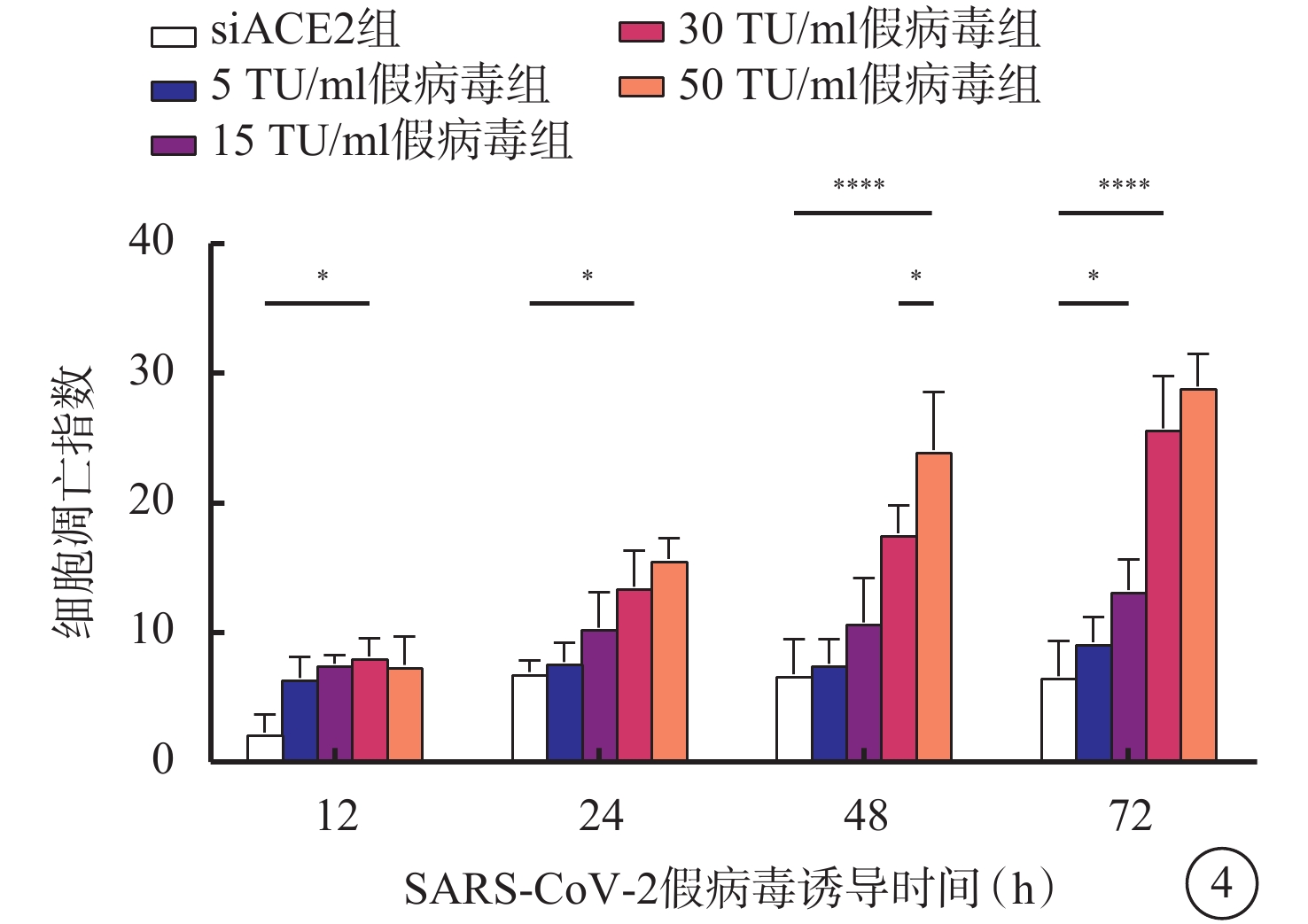
與siACE2組比較,感染組細胞凋亡指數隨假病毒誘導時間延長逐漸升高,差異均有統計學意義(F=2.523、6.716、3.477、3.421,P<0.05、0.05、0.01、0.01)。與siACE2組比較,5 TU/ml假病毒組、15 TU/ml假病毒組細胞誘導12、24、48、72 h后,細胞凋亡指數上調,差異無統計學意義(F=9.185、6.352、6.268、8.419,P>0.05);與siACE2組比較,30 TU/ml假病毒組、50 TU/ml假病毒組細胞凋亡指數顯著升高,差異有統計學意義(F=3.169、6.575、12.440、14.870,P<0.05、0.05、0.01、0.01);而30 TU/ml假病毒組與50 TU/ml假病毒組誘導12、24、72 h比較,無顯著差異(t=0.277、0.9363、2.769,P>0.05),但誘導48 h后,50 TU/ml假病毒組較30 TU/ml假病毒組細胞存活率明顯降低,差異有統計學意義(t=1.394,P<0.05)(圖4)。
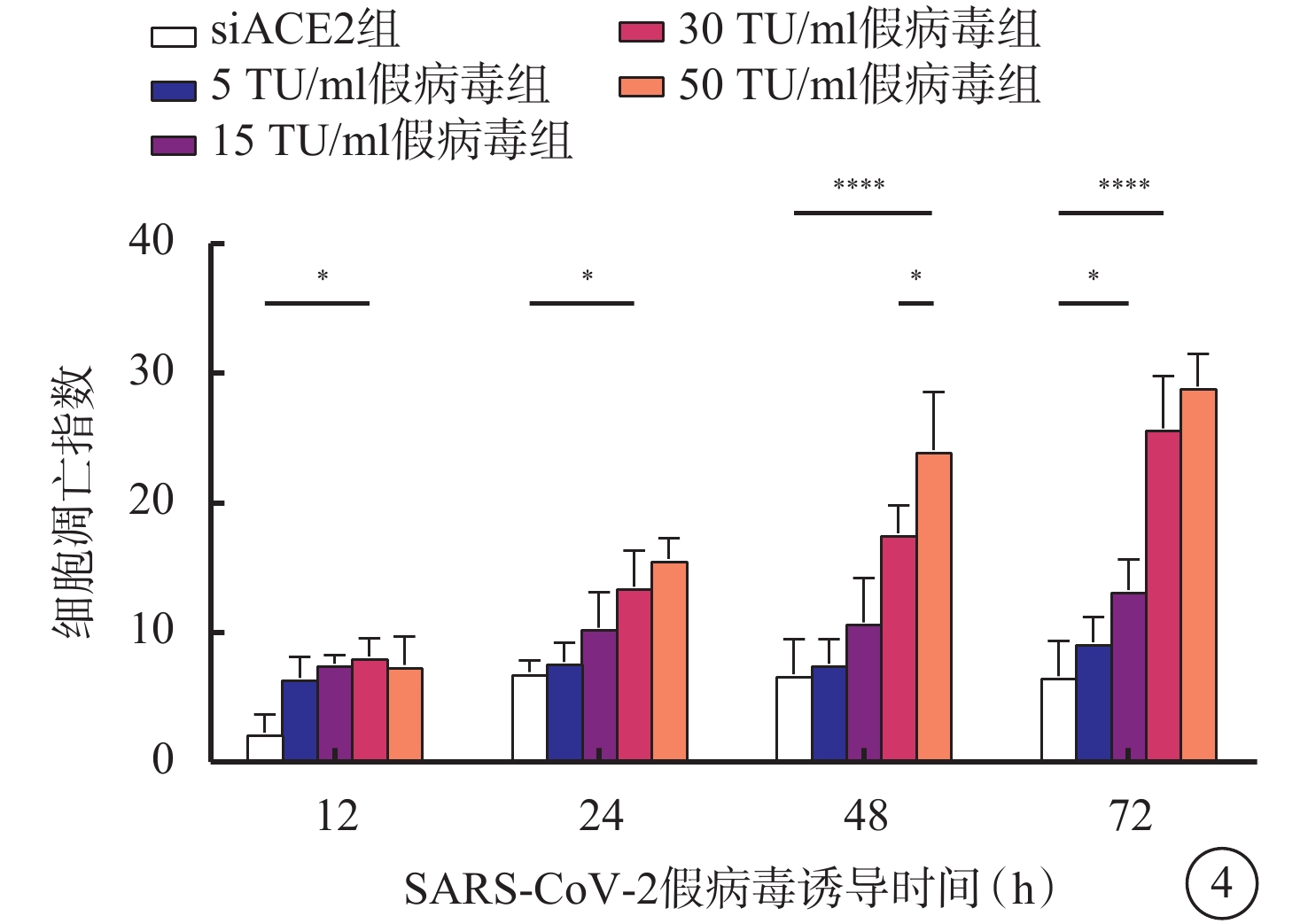 圖4
SARS-CoV-2假病毒不同誘導時間siACE2組與感染組細胞凋亡指數比較(n=3) *P<0.05,****P<0.01 661w細胞:小鼠光感受器細胞;SARS-CoV-2:急性呼吸綜合征冠狀病毒2型;ACE2:血管緊張素轉化酶2;siACE2:ACE2 siRNA過表達慢病毒;siACE2組:661w細胞轉染siACE2;感染組:661w轉染siACE2后,再分別加入5、15、30、50 TU/ml SARS-CoV-2假病毒誘導
圖4
SARS-CoV-2假病毒不同誘導時間siACE2組與感染組細胞凋亡指數比較(n=3) *P<0.05,****P<0.01 661w細胞:小鼠光感受器細胞;SARS-CoV-2:急性呼吸綜合征冠狀病毒2型;ACE2:血管緊張素轉化酶2;siACE2:ACE2 siRNA過表達慢病毒;siACE2組:661w細胞轉染siACE2;感染組:661w轉染siACE2后,再分別加入5、15、30、50 TU/ml SARS-CoV-2假病毒誘導
2.5 假病毒誘導使661w細胞周期阻滯于G1期
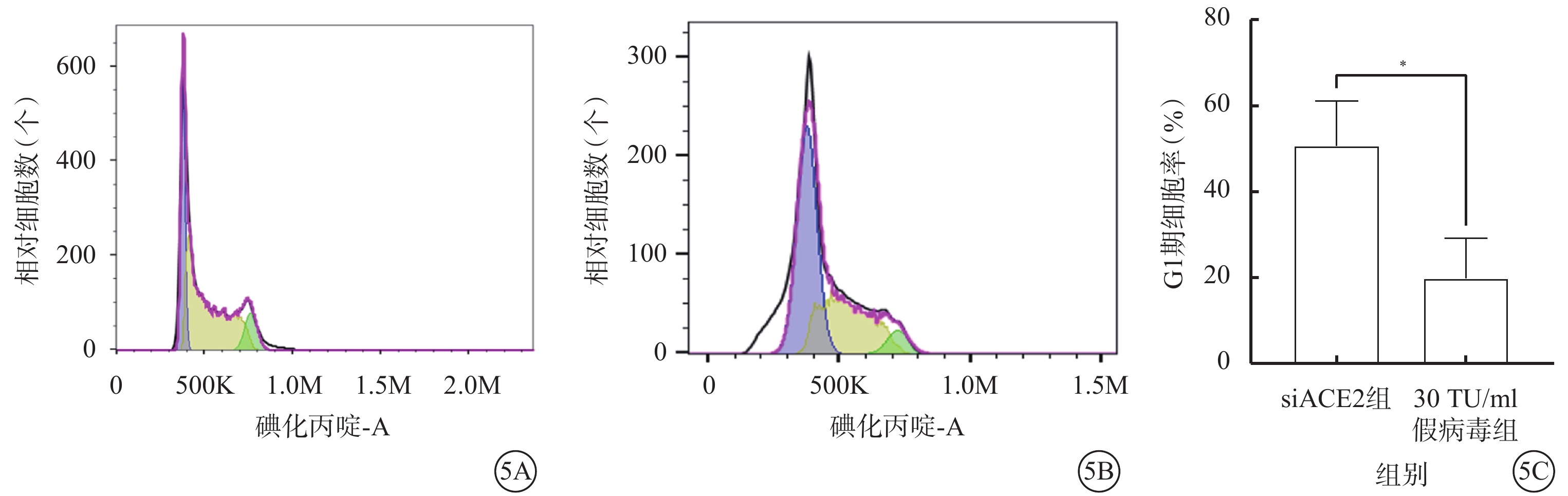
假病毒誘導72 h時,與siACE2組比較,30 TU/ml假病毒組G1期細胞顯著增加,差異有統計學意義(t=3.812,P<0.05)(圖5)。
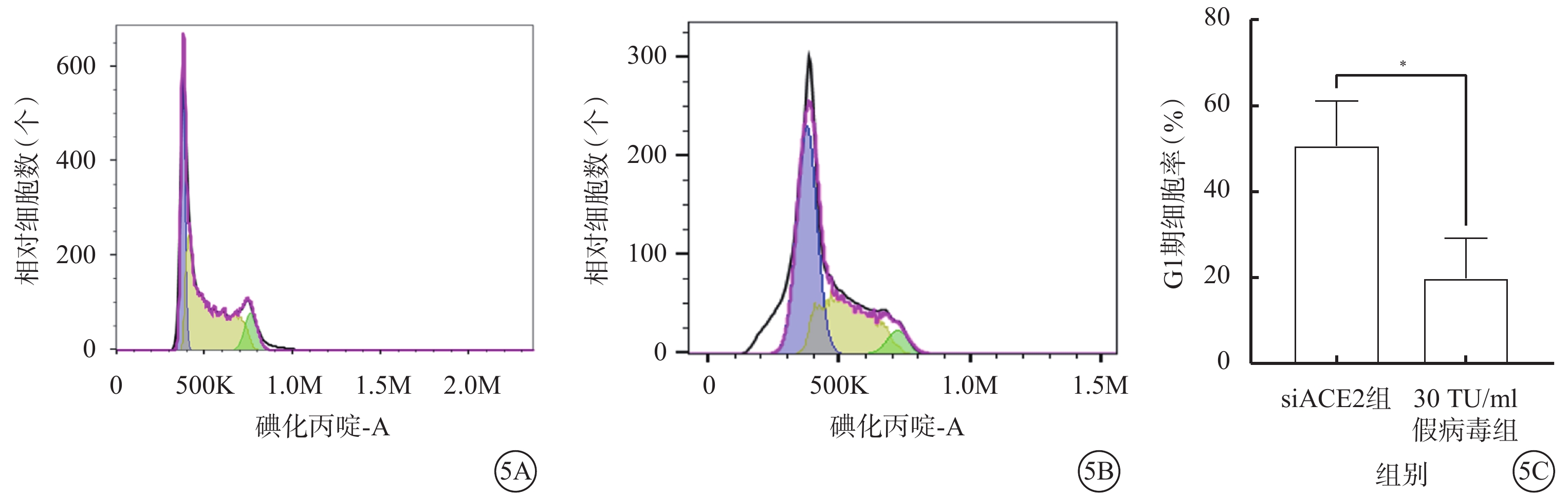 圖5
SARS-CoV-2假病毒誘導72 h促使661w細胞周期阻滯于G1期(n=3) 5A、5B分別示siACE組、30 TU/ml假病毒組流式細胞儀檢測像;5C示siACE2組、30 TU/ml假病毒組細胞周期G1期比較,P<0.05 661w細胞:小鼠光感受器細胞;SARS-CoV-2:急性呼吸綜合征冠狀病毒2型;ACE2:血管緊張素轉化酶2;siACE2:ACE2 siRNA過表達慢病毒;siACE2組:661w細胞轉染siACE2;30 TU/ml假病毒組:661w轉染siACE2后,加入30 TU/ml SARS-CoV-2假病毒誘導
圖5
SARS-CoV-2假病毒誘導72 h促使661w細胞周期阻滯于G1期(n=3) 5A、5B分別示siACE組、30 TU/ml假病毒組流式細胞儀檢測像;5C示siACE2組、30 TU/ml假病毒組細胞周期G1期比較,P<0.05 661w細胞:小鼠光感受器細胞;SARS-CoV-2:急性呼吸綜合征冠狀病毒2型;ACE2:血管緊張素轉化酶2;siACE2:ACE2 siRNA過表達慢病毒;siACE2組:661w細胞轉染siACE2;30 TU/ml假病毒組:661w轉染siACE2后,加入30 TU/ml SARS-CoV-2假病毒誘導
2.6 SARS-CoV-2上調661w細胞免疫相關因子、誘導調節凋亡相關因子蛋白及mRNA表達水平
qPCR、Western blot檢測結果顯示,假病毒誘導72 h時,與siACE2組比較,30 TU/ml假病毒組細胞中IL-6、TNF-α、Bax蛋白和mRNA相對表達量均上調(t=7.601、6.039、3.088、5.193、6.427、7.667),Bcl-2蛋白和mRNA相對表達量下調(t=3.614、6.777),差異均有統計學意義(P<0.05)(圖6)。
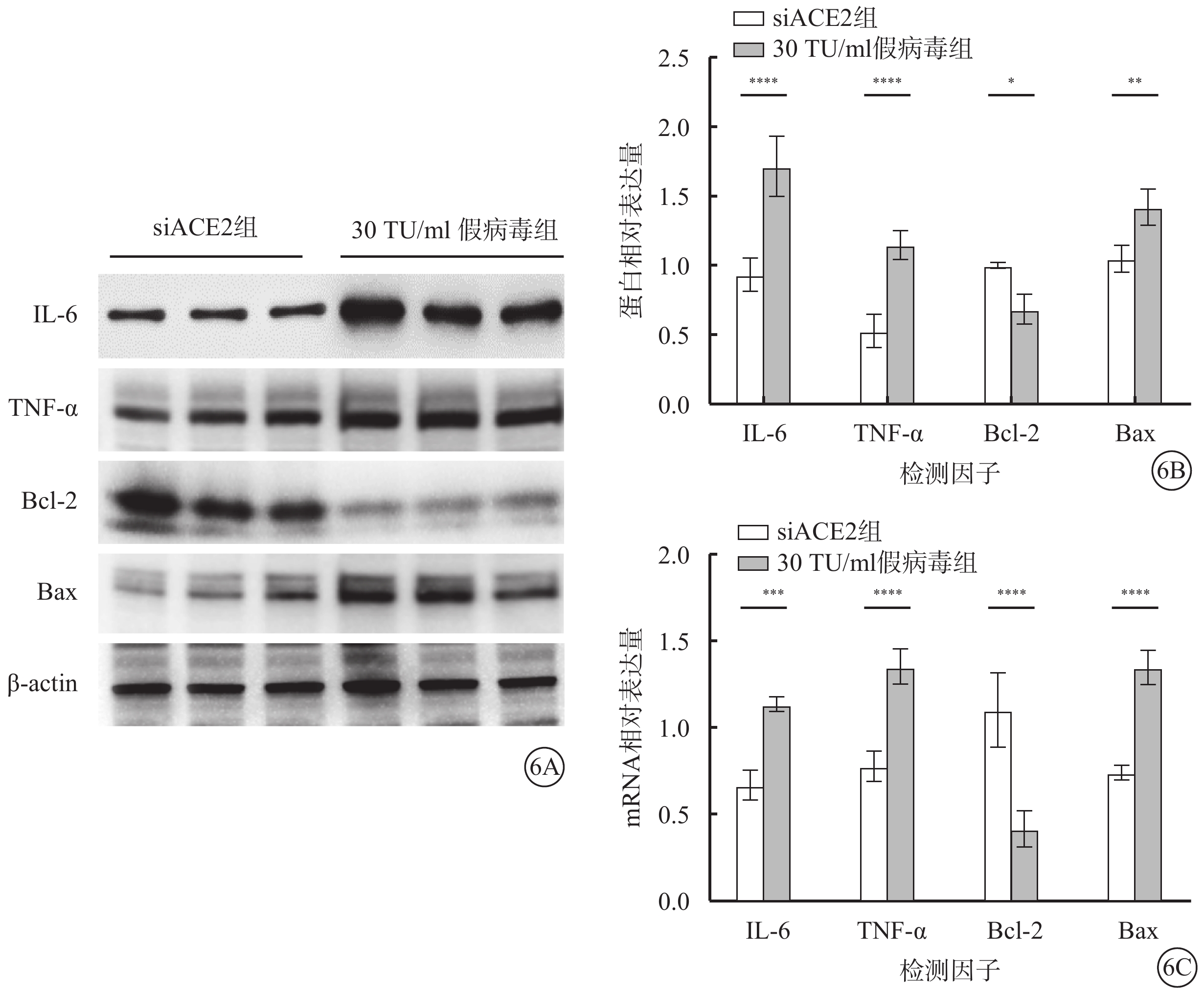 圖6
SARS-CoV-2假病毒誘導72 h促進661w細胞中IL-6、TNF-α、Bcl-2、Bax蛋白和mRNA表達水平(n=3) 6A示電泳圖;6B、6C分別示siACE組、30 TU/ml假病毒組細胞中IL-6、TNF-α、Bcl-2、Bax蛋白、mRNA相對表達量比較,*P<0.05,**P<0.01 661w細胞:小鼠光感受器細胞;SARS-CoV-2:急性呼吸綜合征冠狀病毒2型;ACE2:血管緊張素轉化酶2;siACE2:ACE2 siRNA過表達慢病毒;siACE2組:661w細胞轉染siACE2;30 TU/ml假病毒組:661w細胞轉染siACE2后,加入30 TU/ml SARS-CoV-2假病毒誘導;IL-6:白細胞介素-6;TNF-α:腫瘤壞死因子-α;Bcl-2:B淋巴細胞瘤-2;Bax:Bcl-2相關X蛋白
圖6
SARS-CoV-2假病毒誘導72 h促進661w細胞中IL-6、TNF-α、Bcl-2、Bax蛋白和mRNA表達水平(n=3) 6A示電泳圖;6B、6C分別示siACE組、30 TU/ml假病毒組細胞中IL-6、TNF-α、Bcl-2、Bax蛋白、mRNA相對表達量比較,*P<0.05,**P<0.01 661w細胞:小鼠光感受器細胞;SARS-CoV-2:急性呼吸綜合征冠狀病毒2型;ACE2:血管緊張素轉化酶2;siACE2:ACE2 siRNA過表達慢病毒;siACE2組:661w細胞轉染siACE2;30 TU/ml假病毒組:661w細胞轉染siACE2后,加入30 TU/ml SARS-CoV-2假病毒誘導;IL-6:白細胞介素-6;TNF-α:腫瘤壞死因子-α;Bcl-2:B淋巴細胞瘤-2;Bax:Bcl-2相關X蛋白
2.7 SARS-CoV-2假病毒誘導661w細胞NF-κB1、NF-κB2、P65、P100 mRNA的表達水平
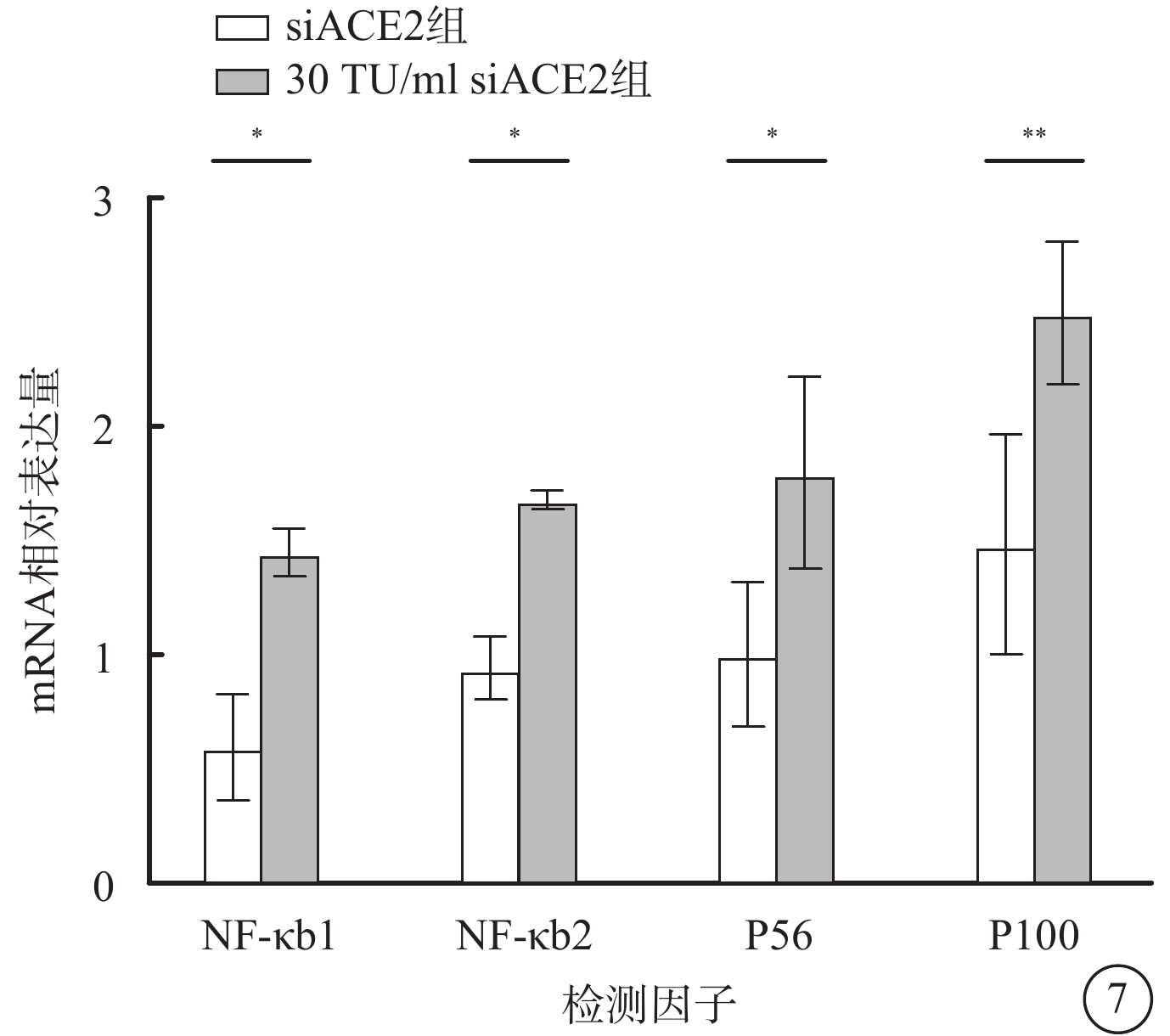
qPCR檢測結果顯示,假病毒誘導72 h時,與siACE2組比較,30 TU/ml假病毒組細胞中NF-κB信號通路相關分子NF-κB1、NF-κB2、P65、P100 mRNA相對表達量明顯升高,差異均有統計學意義(t=3.550、3.074、3.307、4.218,P<0.05)(圖7)。
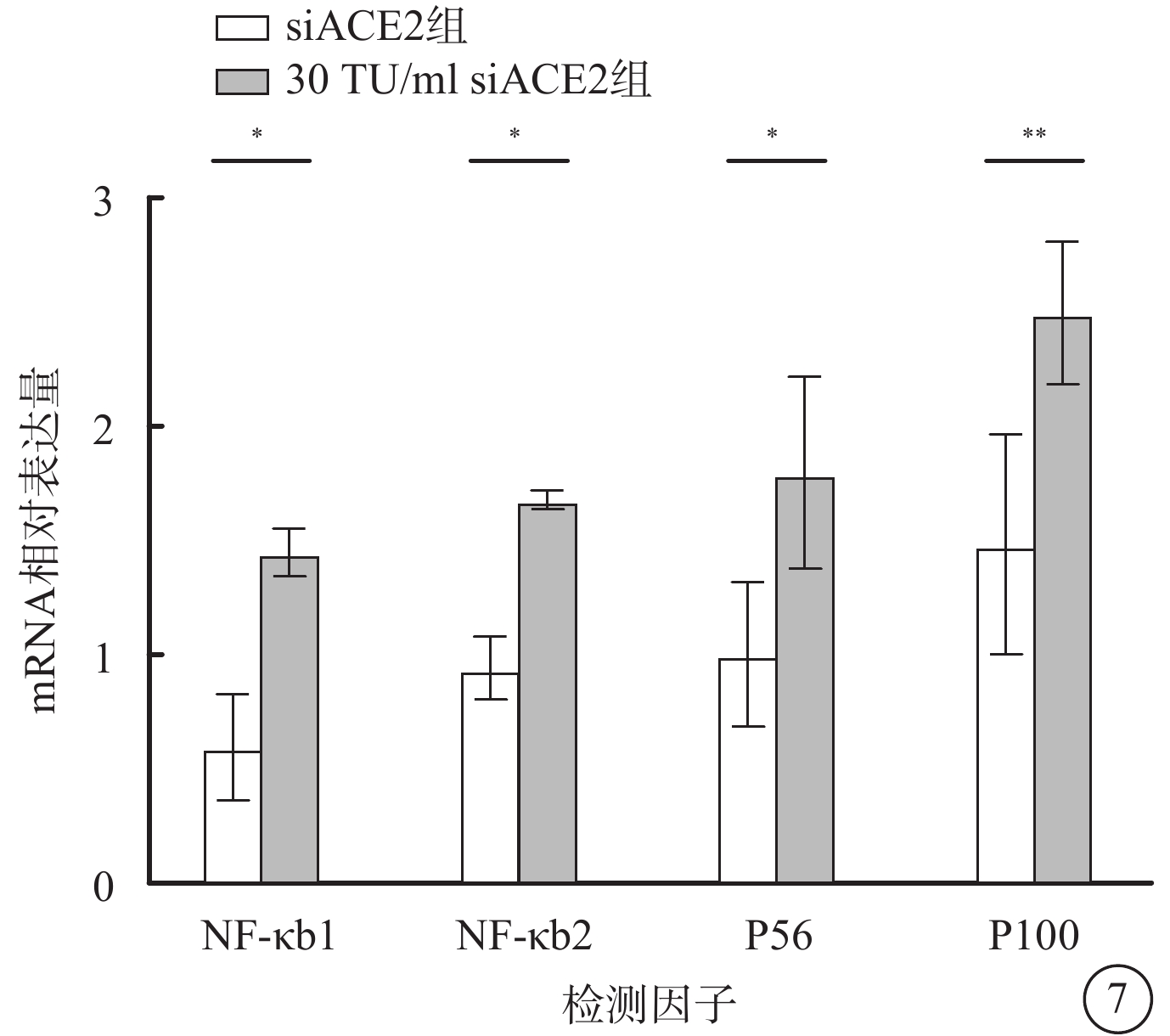 圖7
SARS-CoV-2假病毒誘導72 h siACE2組、30 TU/ml假病毒組661w細胞中NF-κB1、NF-κB2、P65、P100 mRNA相對表達量比較(n=3)*P<0.05,**P<0.01 661w細胞:小鼠光感受器細胞;SARS-CoV-2:急性呼吸綜合征冠狀病毒2型;NF-κB:核因子;P65:NF-κB增強子;P100:NF-κB前體蛋白;ACE2:血管緊張素轉化酶2;siACE2:ACE2 siRNA過表達慢病毒;siACE2組:661w細胞轉染siACE2;30 TU/ml假病毒組:661w細胞轉染siACE2后,加入30 TU/ml SARS-CoV-2假病毒誘導
圖7
SARS-CoV-2假病毒誘導72 h siACE2組、30 TU/ml假病毒組661w細胞中NF-κB1、NF-κB2、P65、P100 mRNA相對表達量比較(n=3)*P<0.05,**P<0.01 661w細胞:小鼠光感受器細胞;SARS-CoV-2:急性呼吸綜合征冠狀病毒2型;NF-κB:核因子;P65:NF-κB增強子;P100:NF-κB前體蛋白;ACE2:血管緊張素轉化酶2;siACE2:ACE2 siRNA過表達慢病毒;siACE2組:661w細胞轉染siACE2;30 TU/ml假病毒組:661w細胞轉染siACE2后,加入30 TU/ml SARS-CoV-2假病毒誘導
3 討論
血管系統是AMN的中心部位,血管系統損傷是AMN患者視覺功能障礙的關鍵[16]。 然而,占視網膜質量和代謝活動大部分的光感受器對疾病的潛在貢獻卻在很大程度上被忽視。研究報道,AMN患者光感受器橢圓體帶發生缺陷,視網膜外核層、光感受器外節變薄,外層視網膜局部出現強反射,并發展為退行性變化[15, 17]。而在COVID-19引起的AMN患者中也發現視網膜光感受器損傷[18]。Menuchin-Lasowski等[19]利用SARS-CoV-2感染視網膜類器官,在其光感受器細胞中優先檢測到SARS-COV-2病毒S蛋白,并誘發光感受器細胞炎癥;使用SARS-CoV-2假病毒對小鼠人源性ACE2(hACE2)光感受器感染2 、5、7 d后,小鼠視網膜中SARS-CoV-2 S蛋白表達水平隨感染時間延長而逐漸升高,且SARS-CoV-2 S蛋白最先出現在視錐、視桿細胞中。
ACE2是SARS-CoV-2進入宿主細胞的重要受體,SARS-CoV-2主要由S蛋白介導,S蛋白與宿主受體ACE2結合進入宿主細胞[3]。因此,為探究SARS-CoV-2病毒感染對光感受器的影響及分子調控機制,本研究建立了可感染SARS-CoV-2假病毒的ACE2過表達小鼠光感受器細胞株,并利用SARS-CoV-2假病毒感染661w細胞。結果顯示,不同濃度感染組的細胞形態隨時間的增加由細長纖維狀的正常形態變為圓形,并出現明顯皺縮;不同濃度感染組細胞的增殖能力較siACE2組顯著降低。有文獻報道,SARS-CoV-2可誘導小膠質細胞及視錐細胞發生凋亡,在視網膜變性進展中發揮重要作用[20]。本研究結果顯示,SARS-CoV-2感染661w細胞后,感染組細胞凋亡指數較siACE2組顯著增加。這表明SARS-CoV-2病毒感染可促進視網膜光感受器細胞發生凋亡。Western blot及qPCR檢測結果證實,30 TU/ml假病毒感染72 h時,661w細胞中促凋亡因子Bax蛋白、mRNA表達水平較siACE2組顯著上調,而抑制凋亡因子Bcl-2蛋白、mRNA表達水平較siACE2組顯著下調。因此,我們推測SARS-CoV-2感染誘導光感受器細胞凋亡可能是AMN的潛在發病機制之一。
研究表明,SARS-CoV-2誘導的過度炎癥反應是患病嚴重程度和死亡的主要原因[21]。COVID-19住院患者血清炎癥因子表達水平,如IL-6、TNF-α、IL-1β顯著升高,其表達程度與疾病嚴重程度呈正相關[21]。有研究應用,SARS-CoV-2感染k18-hACE2轉基因小鼠,發現SARS-CoV-2可破壞血視網膜屏障進而侵入視網膜,并誘導視網膜發生過度炎癥反應,影響視網膜免疫應答功能[22]。因此,為進一步探究SARS-CoV-2感染對光感受器細胞免疫應答功能的影響,本研究分析了SARS-CoV-2假病毒感染的小鼠光感受器細胞中免疫相關因子IL-6、TNF-α的表達,結果顯示,30 TU/ml假病毒感染72 h時,661w細胞中的IL-6、TNF-α的蛋白及mRNA表達水平較siACE2組顯著升高。這提示SARS-CoV-2感染可通過促進光感受器細胞中炎癥因子表達參與AMN進展中的免疫應答反應。
NF-κB信號通路是調控細胞凋亡及免疫應答的重要信號通路之一[23]。既往研究發現,SARS-CoV-2感染會導致細胞發生程序性細胞死亡,并調節NF-κB信號通路,使促炎因子表達水平上調,進而導致細胞免疫應答障礙[24-25]。為進一步探究SARS-CoV-2感染影響光感受器細胞的增殖、凋亡及免疫應答反應的分子調控機制,本研究檢測了SARS-CoV-2感染的光感受器細胞中NF-κB信號通路相關分子的表達情況。結果顯示,30 TU/ml假病毒組感染72 h,661w細胞中NF-κB信號通路相關分子NF-κB1、NF-κB2、P65、P100 mRNA表達水平較siACE2組均顯著升高,提示SARS-CoV-2感染的光感受器細胞中NF-kB通路被激活,推測SARS-CoV-2感染可能通過激活光感受器細胞中NF-κB信號通路,進而抑制光感受器細胞增殖,促進細胞凋亡及免疫相關因子表達。此外,有研究發現,與SARS-CoV-2結合的ACE2受體在人類視覺系統和中樞神經系統中表達,而中樞神經系統的功能障礙可對視網膜功能產生影響[26-27]。通過比較SARS-CoV-2感染后患者的腦部組織和視網膜組織中的基因,發現NF-kB通路被激活并誘導細胞凋亡、免疫相關基因富集到視網膜器官組織中,抑制視網膜中T細胞活化,從而導致視網膜結構或功能發生改變[28-29],這也進一步支持了本研究結果。
SARS-CoV-2感染可能通過激活NF-κB信號通路,抑制光感受器細胞增殖、促進細胞凋亡及阻滯細胞周期,導致視網膜光感受器細胞結構和免疫功能異常,進而誘發AMN發生。本研究旨在為SARS-CoV-2感染誘發AMN發生的病理機制提供理論依據,未來擬進一步利用hACE2轉基因小鼠,探究SARS-CoV-2感染對小鼠視網膜功能及視網膜光感受器的影響,揭示SARS-CoV-2感染后NF-κB信號通路調控光感受器細胞功能的關鍵分子機制,為臨床預防與治療SARS-CoV-2感染引起的AMN奠定理論基礎。
由嚴重急性呼吸綜合征冠狀病毒2型(SARS-CoV-2)引起的新型冠狀病毒感染(COVID-19)是一種嚴重的疾病[1-2]。SARS-COV-2感染后會促進炎癥反應,引起高凝狀態,導致多種全身并發癥,其入侵視網膜可引起視網膜炎癥及其一系列眼部并發癥,如結膜炎、視網膜出血、視網膜靜脈阻塞、急性黃斑神經視網膜病變(AMN)等[3-6]。AMN是一種多發生于年輕、健康女性的較為罕見的視網膜疾病,其主要癥狀為患者眼部驟然出現1個或多個旁中心暗點,視力正常或輕度下降,黃斑區損傷較大的患者,可導致視力暫時或永久性下降[7]。AMN誘發因素包括口服避孕藥、呼吸系統損傷、流感樣疾病和咖啡因攝入等[8-9]。SARS-CoV-2感染和疫苗接種對AMN發生和發展可能具有一定影響[10-12]。SARS-CoV-2通過尖峰(S)蛋白與其受體血管緊張素轉化酶2(ACE2)結合的方式侵入眼部[13-14]。AMN患者視網膜敏感度降低的區域內,光感受器結構優先破壞,提示視網膜光感受器損傷與AMN發病機制相關[15]。然而,目前關于SARS-CoV-2在視網膜光感受器的研究鮮見報道。本研究通過體外細胞實驗分析SARS-CoV-2對小鼠光感受器細胞(661w細胞)的形態、增殖、凋亡、周期及免疫功能的作用,并初步探討SARS-CoV-2引發AMN的潛在機制。現將結果報道如下。
1 材料和方法
1.1 材料
小鼠光感受器細胞株(661w細胞,上海通派生物科技有限公司);Dulbecco改良Eagle培養基(DMEM,L540KJ)、4',6-二脒基-2-苯基吲哚(DAPI,C0065)(美國Gibco公司);兔抗小鼠腫瘤壞死因子(TNF)-α多克隆抗體(ab307164)、兔抗小鼠B細胞淋巴瘤-2(Bcl-2)相關X蛋白(Bax)單克隆抗體(ab32503)(英國Abcam公司);兔抗小鼠白細胞介素(IL)-6多克隆抗體、小鼠抗小鼠Bcl-2單克隆抗體、兔抗小鼠β-肌動蛋白(actin)、山羊抗兔免疫球蛋白(Ig)G二抗、山羊抗小鼠IgG二抗(武漢三鷹生物技術有限公司);Alexa Fluor 594山羊抗兔IgG二抗、異硫氰酸熒光素(FITC)標記的鬼筆環肽(40735ES75)、細胞凋亡及周期試劑盒(40302E)(上海翌圣生物科技股份有限公司);兔抗小鼠ACE2單克隆抗體(PA5-47488,美國Thermo Fisher公司);Triton X-100(上海生工生物工程股份有限公司);ACE2 siRNA過表達慢病毒(siACE2)(上海吉凱公司);SARS-CoV-2假病毒(PSV005,北京依翹神州生物技術公司);細胞計數試劑盒8(CCK-8,美國Selleck公司);磷酸鹽緩沖液(PBS,北京蘭杰柯科技有限公司);DMEM高糖完全培養基、胰蛋白酶、20倍洗膜緩沖液(TBST)(北京索萊寶科技有限公司);酶聯免疫檢測儀(ELX800,美國Bio-Teck公司)。正置光學顯微鏡(日本Olympus公司);iQ5實時熒光定量聚合酶鏈反應(qPCR)儀、細胞流式檢測儀(美國Thermo Fisher公司);FV3000型激光掃描共聚焦顯微鏡(日本Nikon公司)。
1.2 方法
細胞培養和分組。661w細胞株置于含10%胎牛血清和1%雙抗(青霉素/鏈霉素)的DMEM培養基中,于37℃、含5% CO2的細胞培養箱中培養至貼壁,取對數生長期細胞用于實驗。將661w細胞分為三組,分別為正常組(未經任何處理)、siACE2組(過表達ACE2且未感染假病毒)及感染組(過表達ACE2并感染假病毒),其中感染組再分為5 TU/ml假病毒組、15 TU/ml假病毒組、30 TU/ml 假病毒組、50 TU/ml假病毒組,分別誘導12、24、48、72 h。
構建siACE2過表達光感受器細胞。應用DMEM高糖完全培養基培養661w細胞,在37℃且含5% CO2的細胞培養箱中培養至貼壁。將細胞以1×104個/ml的細胞密度接種于24孔板中,細胞貼壁后,加入1×108 TU/ml siACE2轉染8 h,更換完全培養基,繼續培養48~72 h,細胞密度達到70%~80%時,0.25%不含乙二胺四乙酸(EDTA)胰酶消化細胞,以離心半徑15cm,1 200 r/min離心5 min,完全培養基重懸細胞,以1×105個/ml的661w細胞密度接種于12孔板中,培養至貼壁后,更換含有3 μg/ml Puromycin的DMED完全培養基繼續培養48 h,將ACE2過表達的661w細胞進行擴增及凍存保種。
激光共聚焦顯微鏡檢測ACE2轉染效率。將siACE2轉染后的細胞以5×103個/ml的細胞密度接種于共聚焦細胞培養皿中培養;細胞培養至50%后,4%多聚甲醛固定30 min,PBS漂洗3次,10 min/次;0.5% Triton X-100中孵育30 min,2%牛血清白蛋白封閉,室溫靜置1 h。加入兔抗小鼠ACE2一抗(1∶500),4 ℃過夜孵育,PBS漂洗3次;加入羊抗小鼠IgG抗體(1∶1 000),室溫孵育2 h,PBS漂洗3次,DAPI封片液封片,激光共聚焦顯微鏡下觀察拍照。
光學顯微鏡觀察各組細胞形態變化。將細胞以1×105個/ml的細胞密度接種于6孔板中,培養至細胞貼壁后,分別用假病毒誘導12、24、48、72 h,光學顯微鏡下觀察細胞形態并采集圖像。
CCK-8法檢測siACE組及感染組細胞活性。各組細胞以1×104個/ml的細胞密度接種于96孔板中,分別用假病毒誘導12、24、48、72 h,每孔滴加10 μl CCK-8試劑孵育1 h,酶鏈免疫檢測儀測量波長450 nm處吸光度[A,舊稱光密度(OD)]值,每組設3個復孔,實驗重復3次。計算不同病毒感染濃度及不同感染時間的細胞存活率,統計細胞增殖情況。細胞存活率=[A(感染)-A(空白)/A(對照)-A(空白)]×100%。
流式細胞儀檢測siACE組及感染組細胞凋亡情況。各組細胞以5×104個/ml的細胞密度接種于24孔板中,細胞貼壁后對各組細胞應用假病毒誘導12、24、48、72 h。0.25%無EDTA胰蛋白酶消化細胞,300 ×g離心5 min,加入膜聯蛋白(Annexin) V-FITC探針混勻,室溫避光孵育20min,加入碘化丙啶(PI)染色液混勻,冰浴避光孵育15 min。流式細胞儀檢測細胞凋亡情況,并使用 BDFACSuite軟件進行分析。凋亡指數=(早期凋亡細胞數量+晚期凋亡細胞數量)/總細胞數量。
流式細胞儀檢測siACE組、30 TU/ml假病毒組細胞周期。siACE組、30 TU/ml假病毒組細胞以1×105個/ml的細胞密度接種于24孔板中,假病毒誘導72 h,0.25%胰蛋白酶消化,加入預冷70%乙醇重懸細胞,4 ℃甲醛固定過夜,加入10 μl RNase A、10 μl PI,37°C下避光孵育30 min,流式細胞儀檢測細胞周期。
蛋白質免疫印跡法(Western blot)檢測siACE組、30 TU/ml假病毒組IL-6、TNF-α、Bcl-2、Bax蛋白相對表達量。siACE組、30 TU/ml假病毒組細胞以1×105個/ml的細胞密度接種于6孔板中,假病毒誘導72 h,提取各組細胞總蛋白,調整蛋白濃度。每個樣孔加入30~50 μg待測樣品,十二烷基硫酸鈉聚丙烯酰胺凝膠電泳中上樣電泳;80 V恒壓0.5 h,樣品進入分離膠后100 V恒壓2 h,300 mA轉膜2 h,5%脫脂牛奶常溫封閉2 h,一抗IL-6、TNF-α、Bax、Bcl-2、ACE2、β-actin(1∶1 000) 4℃過夜孵育(β-actin作為陽性對照),TBST洗膜10 min,重復3次;加入辣根過氧化物偶聯的二抗(1∶5 000),37℃孵育2 h,TBST洗膜10 min,重復5次;加入化學發光劑,暗室顯影成像。采用Image J Program軟件分析蛋白條帶灰度值,以目的條帶與β-actin 條帶的比值代表蛋白的相對表達水平。
qPCR檢測siACE組、30 TU/ml假病毒組細胞中IL-6、TNF-α、Bcl-2、Bax、核因子(NF)κB-1、NF-κB2以及NF- κB增強子(P65)、前體蛋白(P100)mRNA相對表達量。siACE組、30 TU/ml假病毒組細胞假病毒感染72 h,加入Trizol溶液裂解細胞;加入1/5總體積氯仿,充分混合并離心;將上層水相移至新管并加入等體積的異丙醇溶液,以離心半徑15 cm,12 000 r/min低溫離心10 min,移除上清液后加入無水乙醇再次低溫離心,倒出上清液,室溫放置干燥30 min,加入20 μl RNAse-free水,吹打溶解,提取各組細胞RNA,酶標儀檢測RNA濃度及純度;384孔板中依此加入2.5 μl cDNA、1 μl正向引物、1 μl反向引物和4 μl SYBR混合,加樣全程在冰上避光操作。以離心半徑7 cm、12 000 r/min離心10 min。引物序列由擎科生物公司設計(表1)。反應條件:50℃反應2 min,95℃變性10 min(1個循環);95℃變性15 s(40個循環);55℃擴增30 s;95℃反應15 s;60℃反應15 s,95℃ 反應15 s。置于qPCR儀中進行擴增并輸出循環閾值(Ct值),以β-actin為內參,依照公式2-ΔΔCt計算mRNA相對表達量。
采用GraphPad Prism v9.1.0.221軟件進行統計學分析。計量數據以均數±標準差(x±s)表示。多組間比較采用單因素方差分析;兩組間比較采用t檢驗。P<0.05為差異有統計學意義
2 結果
2.1 siACE2成功轉染至661w細胞
熒光顯微鏡觀察結果顯示,轉染siACE2 48 h后,661w細胞中ACE2轉染效率>90%(圖1A)。Western blot檢測結果顯示,與正常組比較,siACE組細胞中ACE2蛋白相對表達量顯著增加,差異有統計學意義(t=18.550,P<0.01)(圖1B,1C)。
圖1
siACE2轉染效率驗證(n=3) 1A示661w細胞熒光顯微鏡像(DAPI,標尺:100 μm ),轉染siACE2 48 h后,ACE2轉染效率>90%;1B示電泳圖;1C示正常組、siACE組細胞中ACE2蛋白相對表達量比較, ****P<0.01 661w細胞:小鼠光感受器細胞;actin:肌動蛋白;ACE2:血管緊張素轉化酶2;siACE2:ACE2過表達慢病毒;DAPI:4',6-二脒基-2-苯基吲哚;正常組:661w細胞常規培養;siACE組:661w細胞轉染siACE2
2.2 siACE組及感染組假病毒不同感染時間細胞形態
siACE2組細胞呈細長纖維狀,細胞形態清晰、成簇或平行排列(圖2A~2D);感染組細胞均出現不同程度皺縮(圖2E~2T)。與siACE組比較,5 TU/ml 假病毒組、15 TU/ml假病毒組細胞隨假病毒感染時間增加,細胞形態無明顯改變(圖2E~2L);72 h時,15 TU/ml 假病毒組細胞數量無明顯改變(圖2I~2L),部分細胞形態趨于圓形(圖2L)。30 TU/ml 假病毒組細胞形態隨假病毒誘導時間延長發生明顯改變;12 h、24 h時細胞形態無明顯改變,48 h時部分細胞形態出現皺縮,72 h時細胞嚴重皺縮,形態趨于圓形(圖2M~2P)。50 TU/ml 假病毒組細胞在假病毒誘導24 h時出現皺縮,隨時間延長逐漸嚴重,形態趨于圓形;72 h時細胞數量減少(圖2Q~2T)。
圖2
SARS-CoV-2假病毒不同誘導時間siACE2組與感染組細胞正置光學顯微鏡像(標尺:200 μm) 2A~2D、2E~2H、2I~2L、2M~2P、2Q~2T分別示siACE2組、5 TU/ml假病毒組、15 TU/ml假病毒組、30 TU/ml假病毒組、50 TU/ml假病毒組細胞假病毒誘導12 、24、48、72 h。siACE2組細胞呈細長纖維狀,細胞形態清晰、成簇或平行排列;感染組細胞均出現不同程度皺縮,隨假病毒轉染濃度、時間增加,細胞皺縮加劇,數量減少 661w細胞:小鼠光感受器細胞;SARS-CoV-2:急性呼吸綜合征冠狀病毒2型;ACE2:血管緊張素轉化酶2;siACE2:ACE2 siRNA過表達慢病毒;siACE2組:661w細胞轉染siACE2;感染組:661w細胞轉染siACE2后,分別加入5、15、30、50 TU/ml SARS-CoV-2假病毒誘導
2.3 661w細胞存活率隨假病毒濃度增加、誘導時間延長而降低
與siACE2組比較,感染組低濃度組(5 TU/ml假病毒組、15 TU/ml假病毒組)細胞誘導12、24、48、72 h后細胞存活率出現下調,但差異無統計學意義(F=0.840、0.412、1.498、1.138,P>0.05),感染組高濃度組(30 TU/ml假病毒組、50 TU/ml假病毒組)誘導12、24、48、72 h較siACE2組細胞存活率均降低,差異有統計學意義(F=3.304、4.772、9.629、13.220,P<0.05、0.01、0.01、0.01),而30 TU/ml假病毒組與50 TU/ml假病毒組誘導12、24、72 h比較,無顯著差異(t=3.113、2.030、1.787,P>0.05),但誘導48 h后,50 TU/ml假病毒組較30 TU/ml假病毒組細胞存活率明顯降低,差異有統計學意義(t=4.733,P<0.01)(圖3)。
圖3
SARS-CoV-2假病毒不同誘導時間siACE2組與感染組細胞存活率比較(n=3) *P<0.05,** P<0.01 661w細胞:小鼠光感受器細胞;SARS-CoV-2:急性呼吸綜合征冠狀病毒2型;ACE2:血管緊張素轉化酶2;siACE2:ACE2 siRNA過表達慢病毒;siACE2組:661w細胞轉染siACE2;感染組:661w細胞轉染siACE2后,再分別加入5、15、30、50 TU/ml SARS-CoV-2假病毒誘導
2.4 661w細胞凋亡指數隨SARS-CoV-2假病毒濃度、誘導時間增加而升高
與siACE2組比較,感染組細胞凋亡指數隨假病毒誘導時間延長逐漸升高,差異均有統計學意義(F=2.523、6.716、3.477、3.421,P<0.05、0.05、0.01、0.01)。與siACE2組比較,5 TU/ml假病毒組、15 TU/ml假病毒組細胞誘導12、24、48、72 h后,細胞凋亡指數上調,差異無統計學意義(F=9.185、6.352、6.268、8.419,P>0.05);與siACE2組比較,30 TU/ml假病毒組、50 TU/ml假病毒組細胞凋亡指數顯著升高,差異有統計學意義(F=3.169、6.575、12.440、14.870,P<0.05、0.05、0.01、0.01);而30 TU/ml假病毒組與50 TU/ml假病毒組誘導12、24、72 h比較,無顯著差異(t=0.277、0.9363、2.769,P>0.05),但誘導48 h后,50 TU/ml假病毒組較30 TU/ml假病毒組細胞存活率明顯降低,差異有統計學意義(t=1.394,P<0.05)(圖4)。
圖4
SARS-CoV-2假病毒不同誘導時間siACE2組與感染組細胞凋亡指數比較(n=3) *P<0.05,****P<0.01 661w細胞:小鼠光感受器細胞;SARS-CoV-2:急性呼吸綜合征冠狀病毒2型;ACE2:血管緊張素轉化酶2;siACE2:ACE2 siRNA過表達慢病毒;siACE2組:661w細胞轉染siACE2;感染組:661w轉染siACE2后,再分別加入5、15、30、50 TU/ml SARS-CoV-2假病毒誘導
2.5 假病毒誘導使661w細胞周期阻滯于G1期
假病毒誘導72 h時,與siACE2組比較,30 TU/ml假病毒組G1期細胞顯著增加,差異有統計學意義(t=3.812,P<0.05)(圖5)。
圖5
SARS-CoV-2假病毒誘導72 h促使661w細胞周期阻滯于G1期(n=3) 5A、5B分別示siACE組、30 TU/ml假病毒組流式細胞儀檢測像;5C示siACE2組、30 TU/ml假病毒組細胞周期G1期比較,P<0.05 661w細胞:小鼠光感受器細胞;SARS-CoV-2:急性呼吸綜合征冠狀病毒2型;ACE2:血管緊張素轉化酶2;siACE2:ACE2 siRNA過表達慢病毒;siACE2組:661w細胞轉染siACE2;30 TU/ml假病毒組:661w轉染siACE2后,加入30 TU/ml SARS-CoV-2假病毒誘導
2.6 SARS-CoV-2上調661w細胞免疫相關因子、誘導調節凋亡相關因子蛋白及mRNA表達水平
qPCR、Western blot檢測結果顯示,假病毒誘導72 h時,與siACE2組比較,30 TU/ml假病毒組細胞中IL-6、TNF-α、Bax蛋白和mRNA相對表達量均上調(t=7.601、6.039、3.088、5.193、6.427、7.667),Bcl-2蛋白和mRNA相對表達量下調(t=3.614、6.777),差異均有統計學意義(P<0.05)(圖6)。
圖6
SARS-CoV-2假病毒誘導72 h促進661w細胞中IL-6、TNF-α、Bcl-2、Bax蛋白和mRNA表達水平(n=3) 6A示電泳圖;6B、6C分別示siACE組、30 TU/ml假病毒組細胞中IL-6、TNF-α、Bcl-2、Bax蛋白、mRNA相對表達量比較,*P<0.05,**P<0.01 661w細胞:小鼠光感受器細胞;SARS-CoV-2:急性呼吸綜合征冠狀病毒2型;ACE2:血管緊張素轉化酶2;siACE2:ACE2 siRNA過表達慢病毒;siACE2組:661w細胞轉染siACE2;30 TU/ml假病毒組:661w細胞轉染siACE2后,加入30 TU/ml SARS-CoV-2假病毒誘導;IL-6:白細胞介素-6;TNF-α:腫瘤壞死因子-α;Bcl-2:B淋巴細胞瘤-2;Bax:Bcl-2相關X蛋白
2.7 SARS-CoV-2假病毒誘導661w細胞NF-κB1、NF-κB2、P65、P100 mRNA的表達水平
qPCR檢測結果顯示,假病毒誘導72 h時,與siACE2組比較,30 TU/ml假病毒組細胞中NF-κB信號通路相關分子NF-κB1、NF-κB2、P65、P100 mRNA相對表達量明顯升高,差異均有統計學意義(t=3.550、3.074、3.307、4.218,P<0.05)(圖7)。
圖7
SARS-CoV-2假病毒誘導72 h siACE2組、30 TU/ml假病毒組661w細胞中NF-κB1、NF-κB2、P65、P100 mRNA相對表達量比較(n=3)*P<0.05,**P<0.01 661w細胞:小鼠光感受器細胞;SARS-CoV-2:急性呼吸綜合征冠狀病毒2型;NF-κB:核因子;P65:NF-κB增強子;P100:NF-κB前體蛋白;ACE2:血管緊張素轉化酶2;siACE2:ACE2 siRNA過表達慢病毒;siACE2組:661w細胞轉染siACE2;30 TU/ml假病毒組:661w細胞轉染siACE2后,加入30 TU/ml SARS-CoV-2假病毒誘導
3 討論
血管系統是AMN的中心部位,血管系統損傷是AMN患者視覺功能障礙的關鍵[16]。 然而,占視網膜質量和代謝活動大部分的光感受器對疾病的潛在貢獻卻在很大程度上被忽視。研究報道,AMN患者光感受器橢圓體帶發生缺陷,視網膜外核層、光感受器外節變薄,外層視網膜局部出現強反射,并發展為退行性變化[15, 17]。而在COVID-19引起的AMN患者中也發現視網膜光感受器損傷[18]。Menuchin-Lasowski等[19]利用SARS-CoV-2感染視網膜類器官,在其光感受器細胞中優先檢測到SARS-COV-2病毒S蛋白,并誘發光感受器細胞炎癥;使用SARS-CoV-2假病毒對小鼠人源性ACE2(hACE2)光感受器感染2 、5、7 d后,小鼠視網膜中SARS-CoV-2 S蛋白表達水平隨感染時間延長而逐漸升高,且SARS-CoV-2 S蛋白最先出現在視錐、視桿細胞中。
ACE2是SARS-CoV-2進入宿主細胞的重要受體,SARS-CoV-2主要由S蛋白介導,S蛋白與宿主受體ACE2結合進入宿主細胞[3]。因此,為探究SARS-CoV-2病毒感染對光感受器的影響及分子調控機制,本研究建立了可感染SARS-CoV-2假病毒的ACE2過表達小鼠光感受器細胞株,并利用SARS-CoV-2假病毒感染661w細胞。結果顯示,不同濃度感染組的細胞形態隨時間的增加由細長纖維狀的正常形態變為圓形,并出現明顯皺縮;不同濃度感染組細胞的增殖能力較siACE2組顯著降低。有文獻報道,SARS-CoV-2可誘導小膠質細胞及視錐細胞發生凋亡,在視網膜變性進展中發揮重要作用[20]。本研究結果顯示,SARS-CoV-2感染661w細胞后,感染組細胞凋亡指數較siACE2組顯著增加。這表明SARS-CoV-2病毒感染可促進視網膜光感受器細胞發生凋亡。Western blot及qPCR檢測結果證實,30 TU/ml假病毒感染72 h時,661w細胞中促凋亡因子Bax蛋白、mRNA表達水平較siACE2組顯著上調,而抑制凋亡因子Bcl-2蛋白、mRNA表達水平較siACE2組顯著下調。因此,我們推測SARS-CoV-2感染誘導光感受器細胞凋亡可能是AMN的潛在發病機制之一。
研究表明,SARS-CoV-2誘導的過度炎癥反應是患病嚴重程度和死亡的主要原因[21]。COVID-19住院患者血清炎癥因子表達水平,如IL-6、TNF-α、IL-1β顯著升高,其表達程度與疾病嚴重程度呈正相關[21]。有研究應用,SARS-CoV-2感染k18-hACE2轉基因小鼠,發現SARS-CoV-2可破壞血視網膜屏障進而侵入視網膜,并誘導視網膜發生過度炎癥反應,影響視網膜免疫應答功能[22]。因此,為進一步探究SARS-CoV-2感染對光感受器細胞免疫應答功能的影響,本研究分析了SARS-CoV-2假病毒感染的小鼠光感受器細胞中免疫相關因子IL-6、TNF-α的表達,結果顯示,30 TU/ml假病毒感染72 h時,661w細胞中的IL-6、TNF-α的蛋白及mRNA表達水平較siACE2組顯著升高。這提示SARS-CoV-2感染可通過促進光感受器細胞中炎癥因子表達參與AMN進展中的免疫應答反應。
NF-κB信號通路是調控細胞凋亡及免疫應答的重要信號通路之一[23]。既往研究發現,SARS-CoV-2感染會導致細胞發生程序性細胞死亡,并調節NF-κB信號通路,使促炎因子表達水平上調,進而導致細胞免疫應答障礙[24-25]。為進一步探究SARS-CoV-2感染影響光感受器細胞的增殖、凋亡及免疫應答反應的分子調控機制,本研究檢測了SARS-CoV-2感染的光感受器細胞中NF-κB信號通路相關分子的表達情況。結果顯示,30 TU/ml假病毒組感染72 h,661w細胞中NF-κB信號通路相關分子NF-κB1、NF-κB2、P65、P100 mRNA表達水平較siACE2組均顯著升高,提示SARS-CoV-2感染的光感受器細胞中NF-kB通路被激活,推測SARS-CoV-2感染可能通過激活光感受器細胞中NF-κB信號通路,進而抑制光感受器細胞增殖,促進細胞凋亡及免疫相關因子表達。此外,有研究發現,與SARS-CoV-2結合的ACE2受體在人類視覺系統和中樞神經系統中表達,而中樞神經系統的功能障礙可對視網膜功能產生影響[26-27]。通過比較SARS-CoV-2感染后患者的腦部組織和視網膜組織中的基因,發現NF-kB通路被激活并誘導細胞凋亡、免疫相關基因富集到視網膜器官組織中,抑制視網膜中T細胞活化,從而導致視網膜結構或功能發生改變[28-29],這也進一步支持了本研究結果。
SARS-CoV-2感染可能通過激活NF-κB信號通路,抑制光感受器細胞增殖、促進細胞凋亡及阻滯細胞周期,導致視網膜光感受器細胞結構和免疫功能異常,進而誘發AMN發生。本研究旨在為SARS-CoV-2感染誘發AMN發生的病理機制提供理論依據,未來擬進一步利用hACE2轉基因小鼠,探究SARS-CoV-2感染對小鼠視網膜功能及視網膜光感受器的影響,揭示SARS-CoV-2感染后NF-κB信號通路調控光感受器細胞功能的關鍵分子機制,為臨床預防與治療SARS-CoV-2感染引起的AMN奠定理論基礎。