低強度超聲刺激視網膜具有對初級視覺皮層(V1)的神經活動進行調節的能力,但目前尚不明確不同強度和時間的超聲作用視網膜時,如何調節V1內神經活動。在本文中,記錄了不同超聲強度和不同時間下小鼠V1腦區的局部場電位(LFP)信號,分析了超聲刺激前1 s至刺激后2 s(–1~2 s)對應的LFP幅值,以及delta、theta、alpha、beta、低gamma頻段的功率和樣本熵。實驗結果表明低強度超聲刺激視網膜時,隨著刺激強度的增加,視覺皮層局部場電位峰峰值呈現線性上升趨勢;delta、theta頻段的功率呈現線性上升趨勢,樣本熵呈現線性下降趨勢。隨著刺激時間的增加,視覺皮層局部場電位峰峰值呈現上升趨勢,上升趨勢逐步減弱;delta頻段的功率呈現上升趨勢,樣本熵呈現線性上升趨勢,而theta頻段的樣本熵呈現下降趨勢。結果表明,低強度視網膜超聲刺激對于視覺皮層的神經活動有顯著的調節作用。本研究能幫助理解超聲刺激對視覺系統功能的調控機制,推動個體化多參數調控治療視覺神經退化、視網膜病變等疾病研究的開展。
引用本文: 王倩倩, 袁毅, 閆佳慶. 低強度視網膜超聲刺激對視覺皮層神經活動的調節作用研究. 生物醫學工程學雜志, 2024, 41(6): 1161-1168. doi: 10.7507/1001-5515.202401047 復制
版權信息: ?四川大學華西醫院華西期刊社《生物醫學工程學雜志》版權所有,未經授權不得轉載、改編
0 引言
視網膜退行性疾病使得視覺中樞無法正常接收到視覺信息,嚴重影響著患者的生活質量,增加了家庭和社會的經濟負擔[1]。它造成的視覺障礙經過標準的屈光矯正和藥物治療均無法有效改善其癥狀[2-3]。因此,如何開發治療視網膜退行性疾病的新技術成為了臨床的重要需求。目前常用的人工視網膜假體通常基于電神經刺激[4-6],是治療不同類型視網膜變性的先進技術。然而,這項技術需要將設備放入眼球,高度侵入性的手術不可避免地具有高風險和昂貴的成本。超聲刺激是一種有效且時空精確的激活視網膜的方法,超聲波刺激具有無創、高空間分辨率和高穿透深度優點[7-10],已被證明是一種有前途的無創神經刺激技術。先前研究使用超聲波調節視網膜神經節細胞(retinal ganglion cell,RGC)的放電活動,研究發現,超聲刺激離體小鼠視網膜能夠激活RGC,縮短了ON-RGC和OFF-RGC的光響應潛伏期,同時增強了部分RGC(ON-和OFF-亞型)的光響應并降低了其他RGC的光響應[11-12]。有研究使用離體的蠑螈視網膜來表征超聲波對完整神經回路的影響,發現超聲刺激可快速調節視覺敏感度。還有研究表明在正常視力或視網膜退化的失明大鼠中,對視網膜的直接超聲刺激可以激發上丘和初級視覺皮層(primary visual cortex,V1)[3]神經元活動。
以上研究表明超聲波可以激活健康和退化視網膜中的神經元。具體來說,超聲波可以引起局部神經元反應并在視覺回路中產生響應模式。但是目前關于超聲刺激視覺神經元如何調節視覺皮層的研究卻很少,超聲刺激視網膜時,視覺皮層如何進行信息編碼目前仍不清楚。
探究視覺系統的信息加工機制是神經科學領域的研究熱點[13-15]。人類的視覺系統主要由接收視覺信息的視網膜、傳遞視覺信息的外側膝狀體以及加工視覺信息的視覺皮層組成[16]。隨著神經成像技術的進步以及計算模型的發展,神經編碼和解碼逐漸成為研究視覺機制的主要手段[17-20]。視覺信息解碼旨在解決從皮層表征到視覺信息的逆向推理問題,基本方法為建立模式識別模型,根據記錄到的大腦響應推測出相應的視覺內容,也可以為腦機接口等前沿新興技術的應用提供基礎[19, 21]。因此我們十分有必要在超聲視覺刺激下,對視覺皮層的信息進行解碼。
超聲參數在超聲刺激中起著重要作用。先前的研究中,研究人員使用不同強度和刺激時間的超聲刺激蠕蟲(超聲參數:基頻10 MHz,頻率1 kHz,脈沖重復頻率,占空比50%),研究發現超聲刺激引起蠕蟲逆轉行為的成功率隨著超聲強度(0~1 MPa)的增加呈現近似線性的增加,且隨著刺激時間(50~400 ms)的增加而增加,但是超過閾值后會達到飽和[22]。另外,研究人員使用不同參數的超聲刺激小鼠的運動皮層激發肢體的運動響應,結果表明運動響應的成功率隨著聲強度(持續時間80 ms,頻率500 kHz,刺激強度0~16.8 W/cm2)和聲持續時間(強度4.2 W/cm2,頻率500 kHz,刺激時間0~320 ms)的增加而增加[23]。在我們先前的研究中發現超聲刺激小鼠運動皮層引起的局部場電位(local field potential,LFP)theta頻段的相對功率隨著超聲強度的增加而降低;低gamma頻段的相對功率隨著超聲強度的增加而增加,LFP的熵值隨著超聲強度和刺激時間的增加而減小(基頻500 kHz,脈沖重復頻率1 kHz,占空比40%,刺激強度0.2~1.1 W/cm2,刺激時間100~400 ms)[24]。我們的另一項研究還發現超聲刺激視覺皮層引起的神經元鈣活動的響應時間半高寬隨著刺激時間的增加而增加(刺激強度0.45 MPa,基頻500 kHz,脈沖重復頻率1 kHz,占空比10%,刺激時間300~700 ms)[25]。以上研究表明超聲刺激對神經的調控效果與超聲參數密切相關。但是超聲視覺刺激引起的視覺皮層神經活動是否與超聲參數(強度和刺激時間)具有依賴關系尚不清楚。
為了回答以上兩個問題,我們使用不同強度和刺激時間的超聲刺激小鼠的視網膜,首次探討了超聲視網膜刺激對視覺皮層的調節作用與超聲參數之間的定量關系。它能夠幫助理解超聲刺激對視覺系統功能的調控機制。本研究旨在明確參數選擇規律,以期推動個體化多參數調控治療視覺神經退化、視網膜病變等疾病研究方向的開展。
1 材料和方法
1.1 動物和分組
本實驗中,使用了20只C57BL/6小鼠(均為雄性,體重20~25 g,北京維通利華實驗動物科技有限公司)。所有實驗環節均符合燕山大學動物倫理要求。在實驗開始前7天起至實驗結束期間,均將小鼠飼養在12小時光照/黑暗循環的標準籠具中,按需提供鼠糧和飲用水。并將小鼠隨機分為超聲刺激強度組和超聲刺激時間組,每組10只。對小鼠視網膜進行超聲刺激,記錄V1的LFP。
1.2 動物實驗流程
利用移動式小動物麻醉機(R530,深圳市瑞沃德生命科技有限公司)對小鼠進行麻醉,異氟烷流通速度設置為2%,氧氣流通速度設置為0.5 L/min。利用小鼠適配器(
1.3 超聲刺激系統
超聲刺激系統由任意波形函數發生器1(DG1022U,普源精電科技股份有限公司)、任意波形函數發生器2(AFG
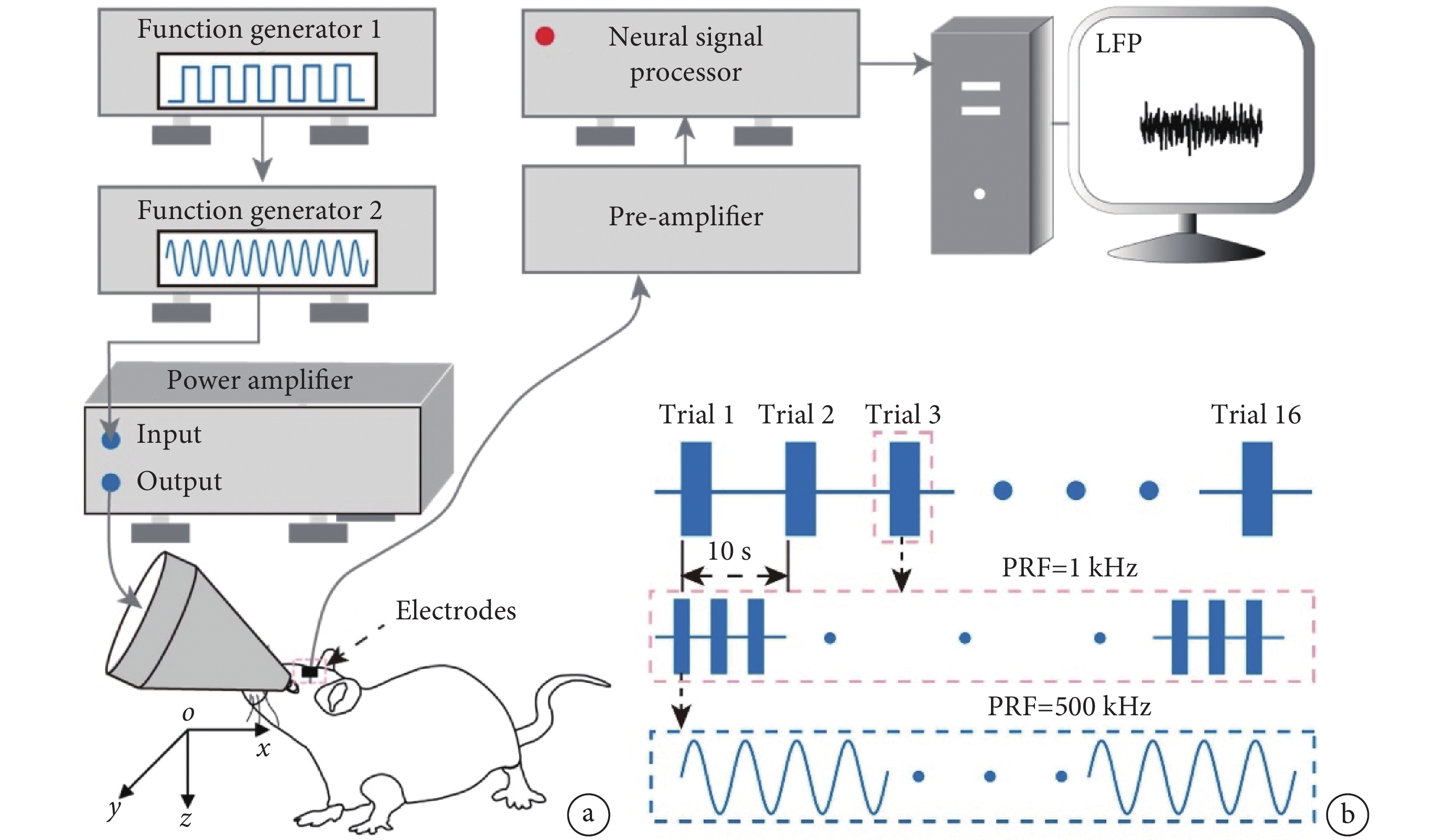
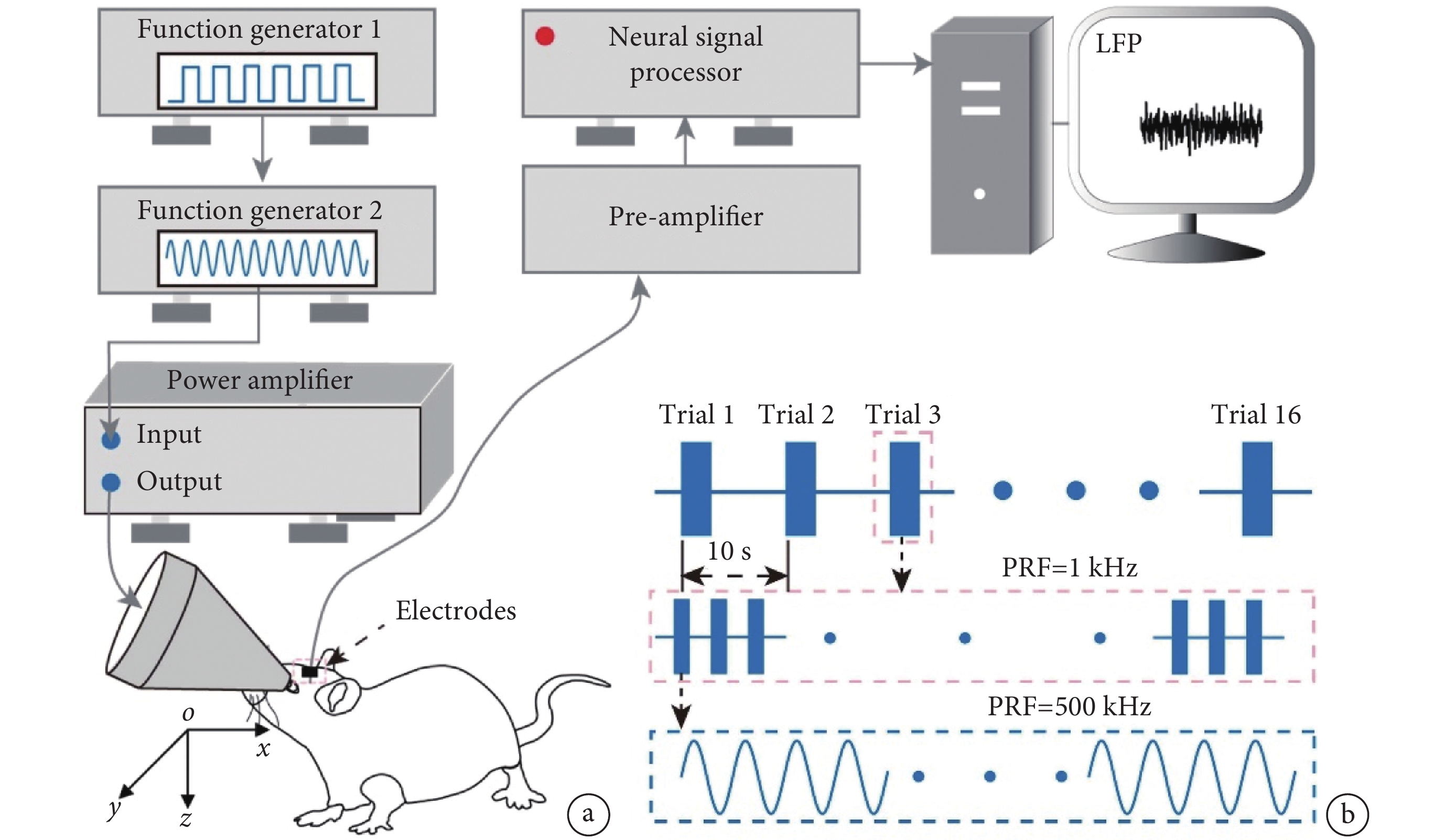 圖1
超聲刺激系統、數據采集系統及超聲刺激參數
圖1
超聲刺激系統、數據采集系統及超聲刺激參數
a. 超聲刺激系統和數據采集系統;b. 超聲刺激參數
Figure1. The ultrasound stimulation system, data acquisition system, and ultrasound stimulation parametersa. ultrasound stimulation system and data acquisition system; b. ultrasound stimulation parameters
1.4 數據采集系統
使用多通道數據采集系統(Apollo,Bio-Signal Technologies,美國)記錄超聲刺激位點和腦電信號。如圖1所示,從小鼠V1采集到的信號經由前置放大器放大,通過采集系統處理得到腦電信號。采集過程中,LFP的采樣率設置為1 kHz。
1.5 LFP預處理
對于鎳鉻電極采集到的16通道LFP信號,去除干擾較大通道的LFP信號,對余下LFP信號進行帶通濾波處理,帶通濾波范圍設置為0.5~200 Hz,并濾掉3 Hz的運動偽跡干擾。
1.6 時頻圖和功率分析
在時頻圖分析過程中采用Welch算法[26],結合刺激位點獲取超聲刺激–1~2 s的LFP數據,分別進行時頻圖處理,對于得到的多通道時頻圖結果進行平均,截取0.5~45 Hz頻段的時頻圖。在功率分析過程中,分別計算了LFP的delta、theta、alpha、beta、低gamma頻段在超聲刺激–1~0、0~1、1~2 s的功率,并觀察不同刺激強度/時間下,各個頻段的腦電功率隨強度/時間變化的情況。
1.7 樣本熵分析
我們參考了先前的文獻[27],分別計算了LFP的delta、theta、alpha、beta、低gamma頻段在超聲刺激–1~0、0~1、1~2 s的樣本熵,并觀察不同刺激強度/時間下,各個頻段的樣本熵隨強度/時間參數變化的情況。
1.8 統計分析
采用多因素方差分析對數據進行統計學分析。檢驗水準為0.05。
2 結果
2.1 不同強度的超聲刺激視網膜時V1的LFP
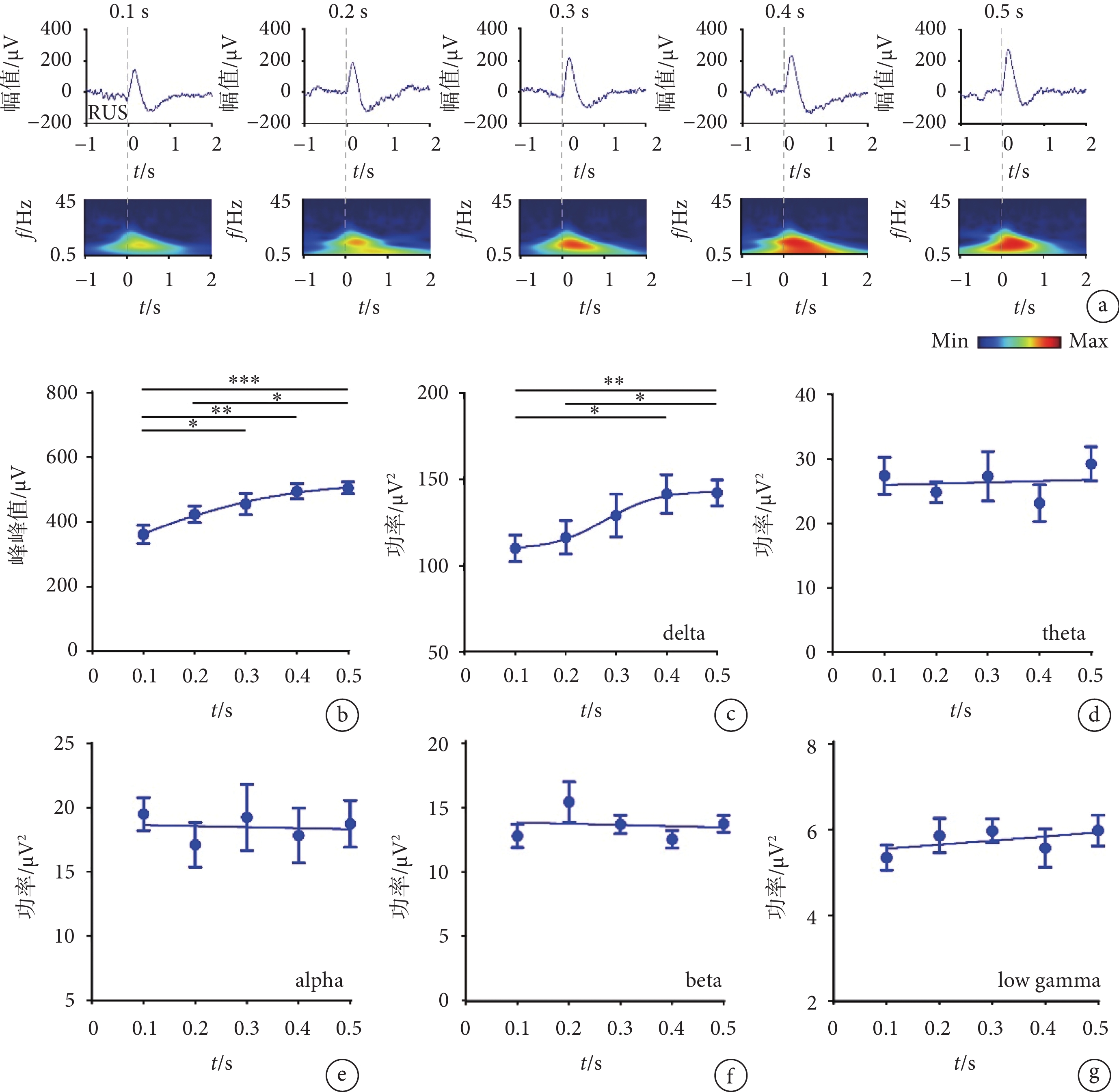
使用不同強度的超聲刺激視網膜,對采集到的V1的LFP進行分析。圖2a中依次呈現了不同刺激強度下(0.1、0.2、0.3、0.4、0.5 MPa)的LFP及時頻圖,可以發現,隨著超聲刺激強度的增加,LFP的幅值呈現上升趨勢,并且在時頻圖的0~1 s,可以觀察到隨著超聲刺激強度的增加,delta頻段的功率有明顯的增強,theta頻段略微增強。如圖2b所示,我們計算了LFP的峰峰值(最高正峰值和最低負峰值的差值),隨著超聲刺激強度的增加,峰峰值呈現明顯的上升趨勢(R2 = 0.694 7)。如圖2c所示,隨著超聲刺激強度的增加,delta頻段的功率同其他頻段相比變化最為明顯,呈現明顯的上升趨勢(R2=0.520 4)。如圖2d所示,隨著超聲刺激強度的增加,theta頻段的功率呈現上升趨勢(R2 =
 圖2
不同強度超聲刺激視網膜時V1的LFP結果
圖2
不同強度超聲刺激視網膜時V1的LFP結果
a. LFP和時頻圖;b. 峰峰值;c-g. delta、theta、alpha、beta、low gamma的功率(mean ± sem,
a. the LFP and time-frequency diagram; b. the peak to peak amplitude; c-g. the power of delta, theta, alpha, beta, and low gamma (mean ± sem,
2.2 不同強度的超聲刺激視網膜時V1的樣本熵
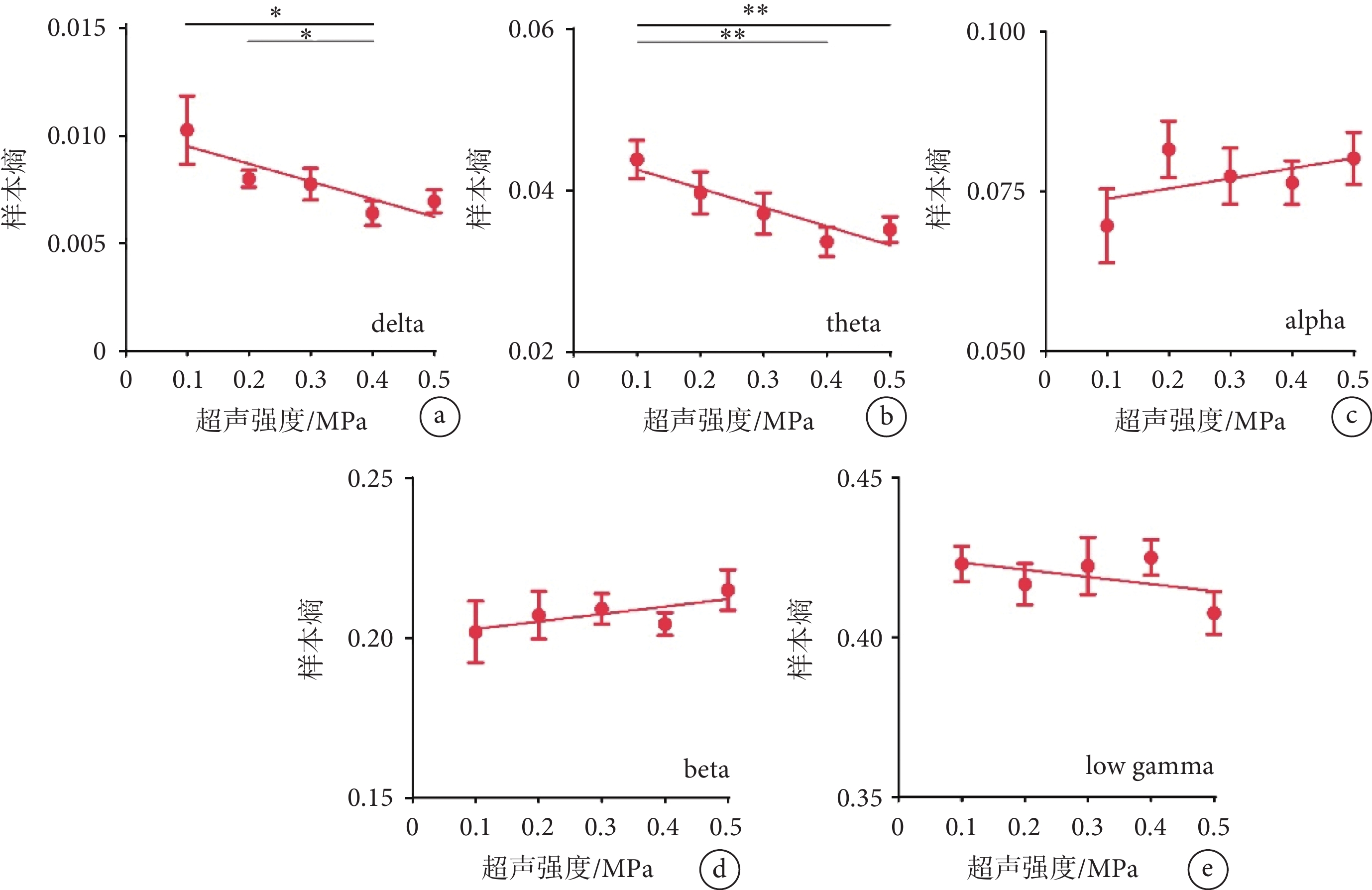
使用不同強度的超聲刺激視網膜時,我們還分析了不同頻段LFP的樣本熵。如圖3a~b所示,隨著超聲刺激強度的增加,delta、theta頻段的樣本熵均呈現下降趨勢(delta:R2 = 0.155 3;theta:R2 = 0.191 3)。如圖3c~e所示,隨著超聲刺激強度的增加,alpha、beta、低gamma頻段的樣本熵無明顯變化。以上結果表明超聲刺激視網膜在V1誘發的LFP低頻段的非線性特征隨著超聲強度的增加而降低。
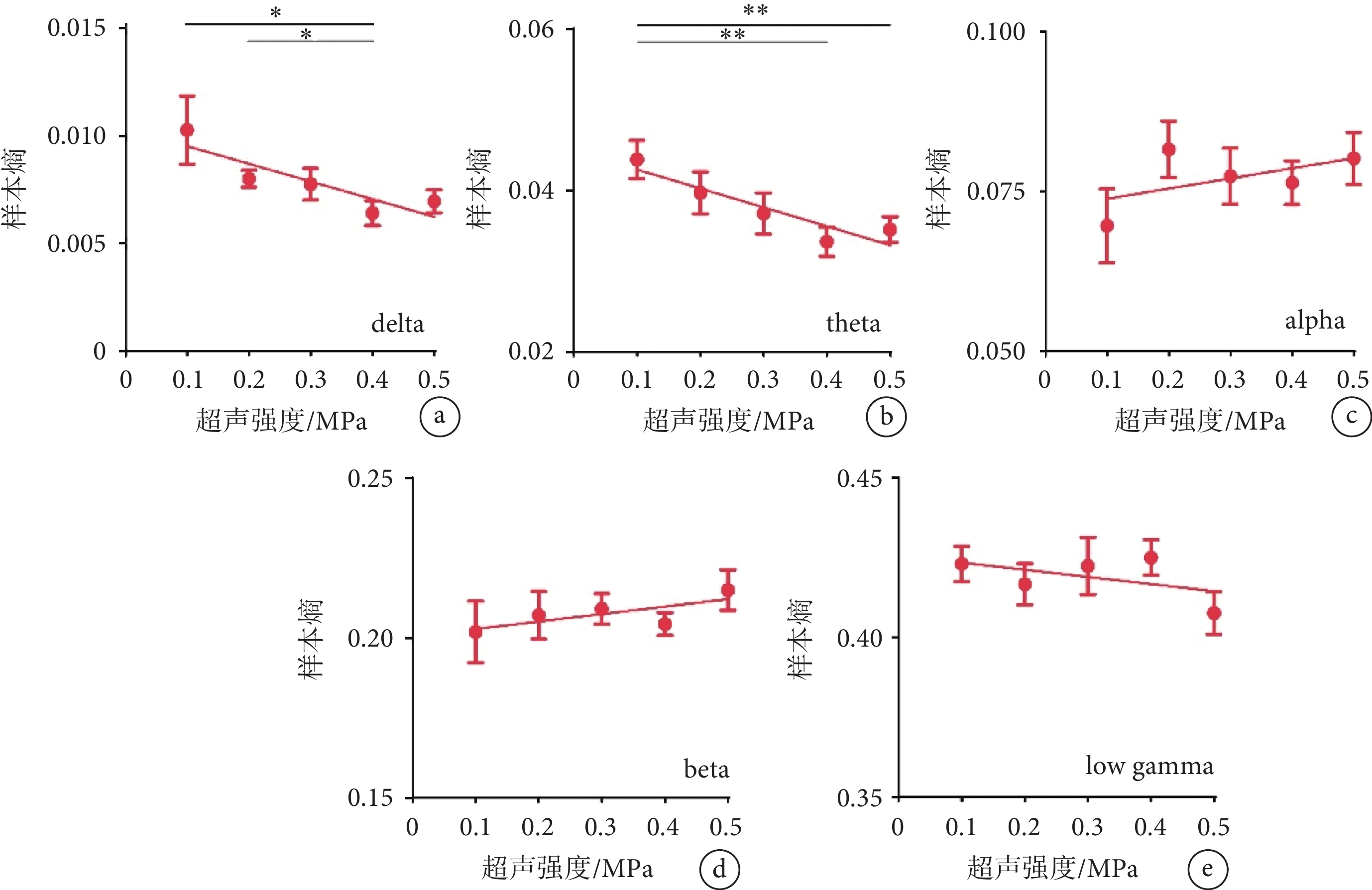 圖3
不同強度超聲刺激視網膜時V1的樣本熵結果
圖3
不同強度超聲刺激視網膜時V1的樣本熵結果
a-e. delta、theta、alpha、beta、low gamma頻段的樣本熵(mean ± sem,
a-e. the sample entropy of delta, theta, alpha, beta, and low gamma (mean ± sem,
2.3 不同時間的超聲刺激視網膜時V1的LFP
使用不同時間的超聲刺激視網膜,對采集到的V1的LFP進行分析。圖4a中依次呈現了不同刺激時間下(0.1、0.2、0.3、0.4、0.5 s)的LFP及時頻圖,可以發現,隨著超聲刺激時間的增加,LFP的幅值也呈現上升趨勢,并且在時頻圖的0~1 s,觀察到delta頻段的功率有明顯增強。如圖4b所示,我們計算了LFP的峰峰值,隨著超聲刺激時間的增加,峰峰值呈現上升趨勢,與不同強度超聲刺激時比較,上升較為緩慢,且隨著超聲刺激時間進一步增加,峰峰值上升趨勢減弱。如圖4c所示,隨著超聲刺激時間的增加,delta頻段的功率整體呈現上升趨勢,刺激時間為0.1~0.3 s時,上升趨勢明顯,0.3 s后上升趨勢開始減弱,0.4~0.5 s之間變化不明顯。如圖4d~g所示,隨著超聲刺激時間的增加,theta、alpha、beta、低gamma頻段的功率無明顯變化。以上結果同樣表明超聲刺激視網膜能夠誘發V1的LFP變化,即引起了神經活動;而且,超聲刺激視網膜誘發的低頻神經活動隨著超聲刺激時間的增加而增加,但是在時間增加到一定程度后一些特征的增加趨勢減弱。
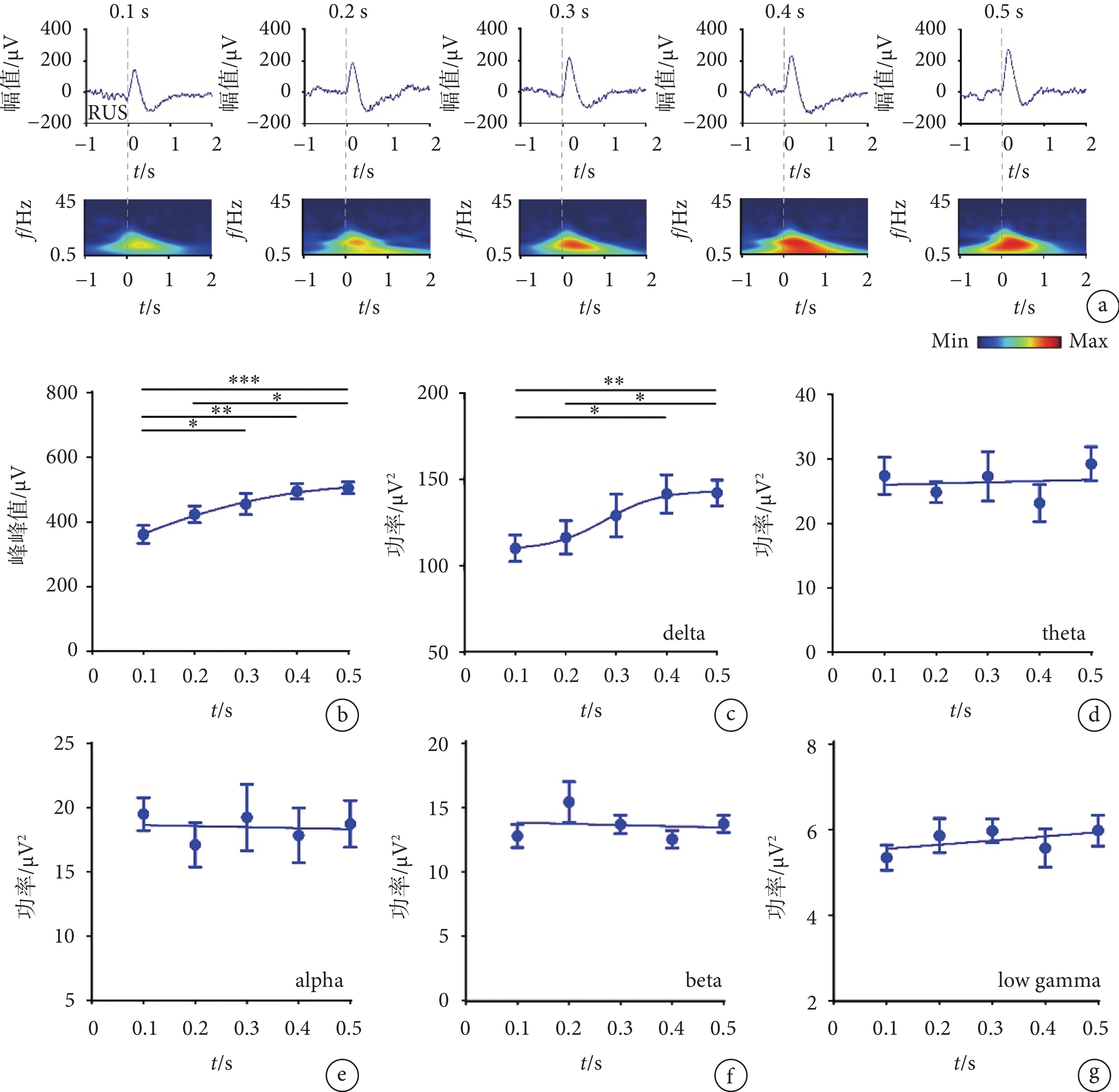 圖4
不同時間超聲刺激視網膜時V1的LFP結果
圖4
不同時間超聲刺激視網膜時V1的LFP結果
a. LFP和時頻圖;b. 峰峰值;c-g. delta、theta、alpha、beta、low gamma的功率(mean ± sem,
a. the LFP and time-frequency diagram; b. the peak to peak amplitude; c-g. the power of delta, theta, alpha, beta, and low gamma (mean ± sem,
2.4 不同時間超聲刺激視網膜時V1的樣本熵
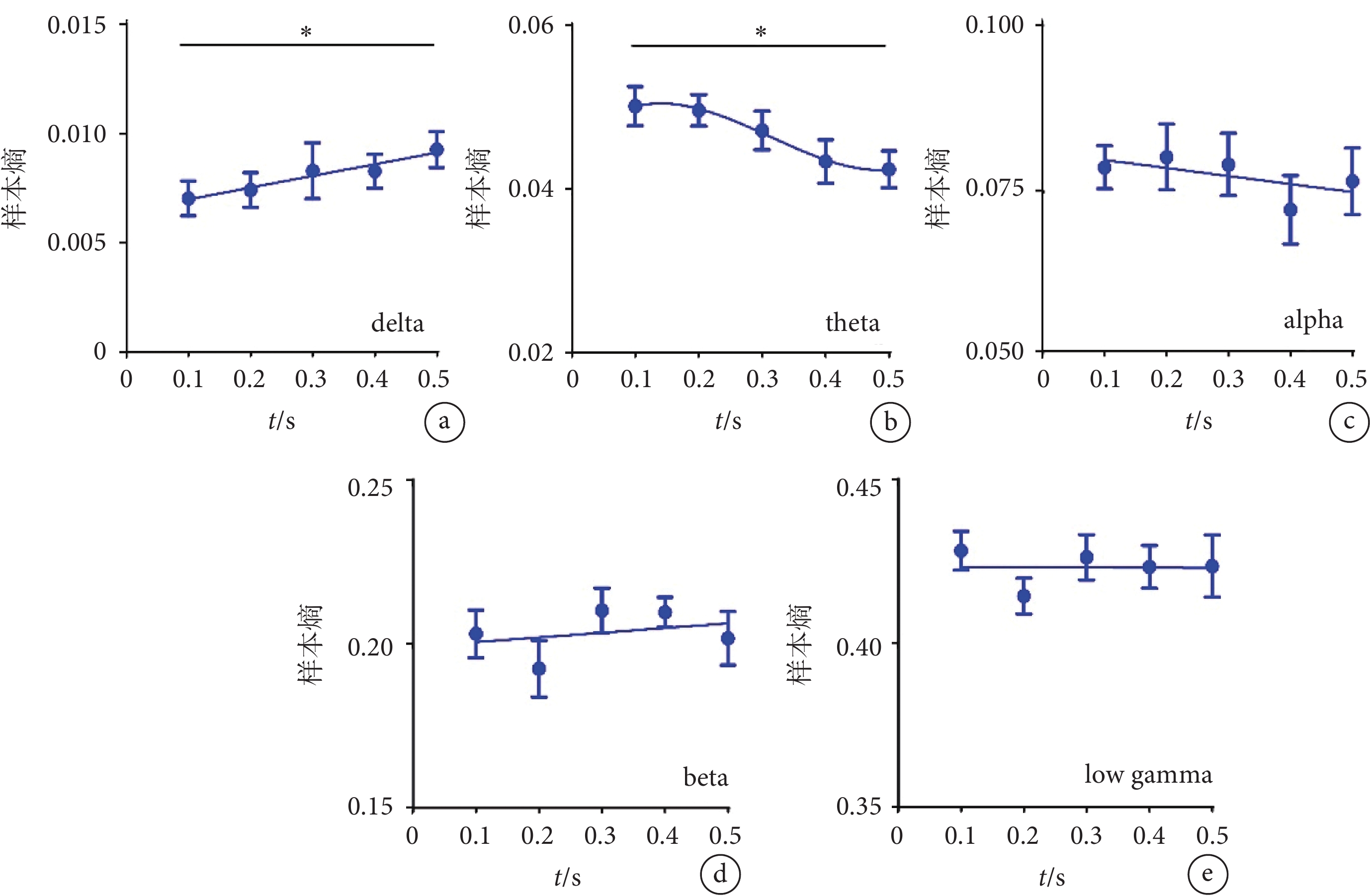
使用不同時間的超聲刺激視網膜時,我們還分析了V1腦區不同頻段LFP的樣本熵。如圖5a所示,隨著超聲刺激時間的增加,delta頻段的樣本熵整體呈現上升趨勢(R2=0.070 3)。如圖5b所示,隨著超聲刺激時間的增加,theta頻段的樣本熵呈現下降趨勢,且隨著超聲刺激時間的進一步增加,下降趨勢減弱。如圖5c~e所示,隨著超聲刺激時間的增加,alpha、beta、低gamma頻段的樣本熵無明顯變化。以上結果表明超聲刺激視網膜可在V1誘發LFP低頻段的非線性特征的變化。該特征在delta頻段隨著超聲刺激時間的增加而升高,在theta頻段隨著超聲刺激時間的增加而降低。
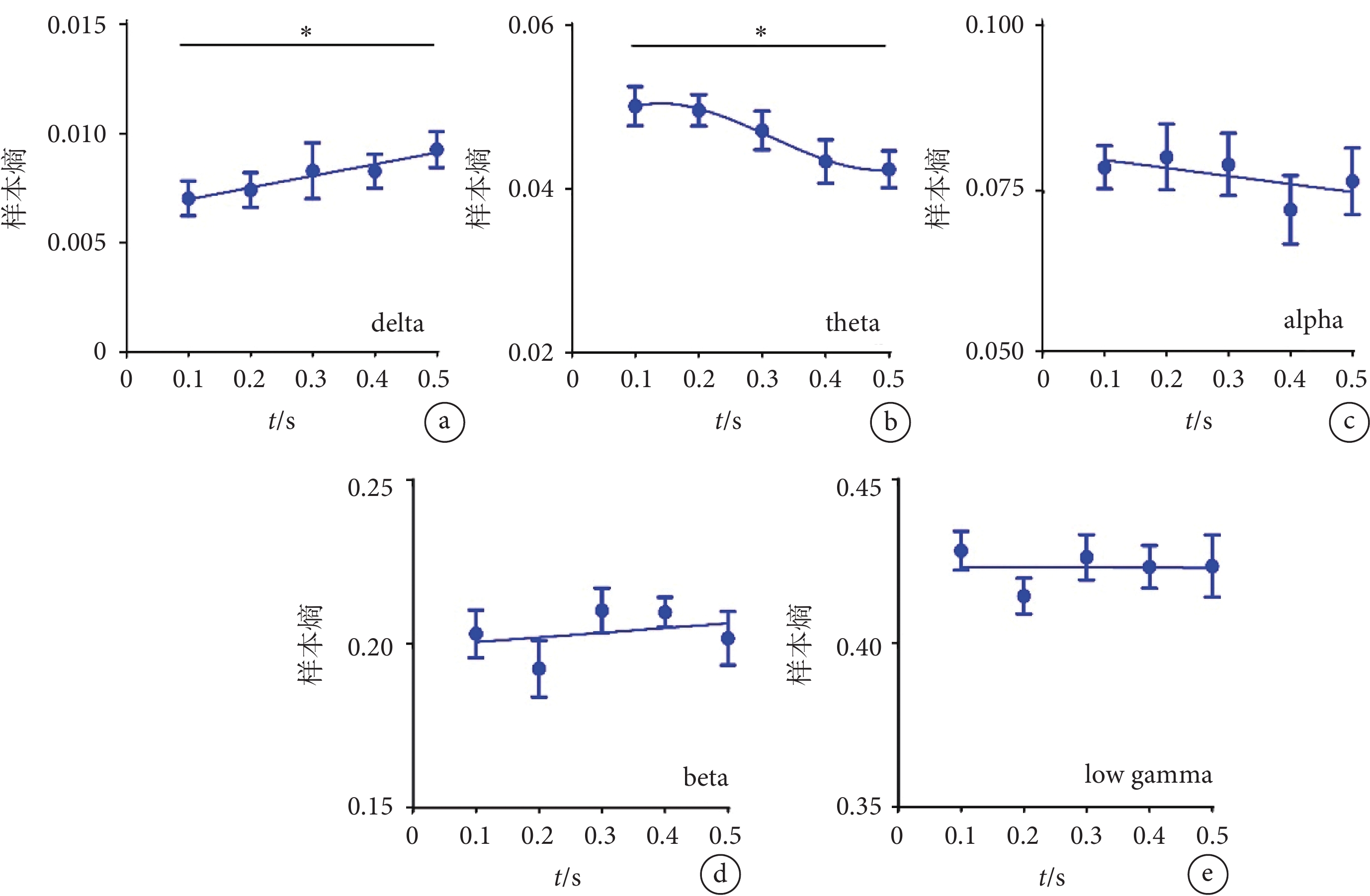 圖5
不同時間超聲刺激視網膜時V1的樣本熵結果
圖5
不同時間超聲刺激視網膜時V1的樣本熵結果
a-e. delta、theta、alpha、beta、low gamma頻段的樣本熵(mean ± sem,
a-e. the sample entropy of delta, theta, alpha, beta, and low gamma (mean ± sem,
3 討論
本文中,我們對小鼠視網膜進行超聲刺激,采集V1的LFP,并對于超聲刺激前后(–1~2 s)的時頻圖、功率及樣本熵進行處理和分析。結果表明,利用不同強度、不同時間的超聲刺激小鼠視網膜時,V1的神經活動會發生改變。隨著超聲強度的增大,V1的LFP峰峰值變大,低頻段的功率增強,非線性特征減弱;隨著超聲刺激時間的增加,V1的LFP峰峰值變大,delta頻段的功率增強但刺激時間增加到一定程度后增強趨勢減弱,delta頻段非線性特征增強,theta頻段非線性特征減弱。以上結果都表明在刺激視網膜時,超聲強度、超聲時間參數的改變都會對V1的神經活動產生調節作用。
我們發現,對于峰峰值、delta頻段功率的變化,超聲刺激強度組的變化趨勢強于超聲刺激時間組。但是何種超聲強度和刺激時間組合對于患者視覺系統的調節作用是最佳的,目前尚不明確,需要結合臨床進一步探討。通過其他學者的研究,我們也注意到如何最優利用超聲刺激,并不局限于超聲參數這一個因素。先前的研究表明,多個視覺區域在功能和視覺感知上是相互聯系的,比如刺激V2或者V3也可以影響V1[28]。視神經、視覺丘腦以及視覺皮層也可以作為研究視覺系統的靶點,已有學者通過視神經內刺激來為全盲患者解決視覺問題[29-30]。在今后的研究中,結合不同的刺激靶點研究不同超聲參數對于視覺系統的調節作用,是一個有價值的研究方向,可為臨床視網膜退化性疾病、視覺損傷疾病的治療提供一定的參考。
先前的研究表明眼部活動和腦部活動之間聯系密切[31],且視網膜神經元可表達諸如大鉀離子(big potassium,BK)、瞬時受體電位(transient receptor potential,TRP)等在內的多個機械敏感離子通道[32]。我們推測低強度超聲刺激可能通過調節視網膜的機械敏感離子通道改變了不同類型神經元之間的分布模式,通過RGC的活動,進而影響神經信號向大腦視覺皮層的傳遞,最終改變視覺皮層的神經活動模式。我們的研究表明,改變超聲刺激強度和時間可以實現對于V1神經活動不同程度的調節。在超聲刺激視網膜改善視覺功能的工作中,希望本研究可以推動多參數協同治療的發展,在未來的研究中可以進一步探索超聲在視網膜退行疾病中的應用潛力,并深入探索視網膜-初級視覺皮層通路可塑性。
重要聲明
利益沖突聲明:本文全體作者均聲明不存在利益沖突。
作者貢獻聲明:王倩倩負責實驗設計、數據采集、數據處理和論文初稿,袁毅負責實驗監督以及論文審查和編輯,閆佳慶負責實驗監督和數據處理。
倫理聲明:本研究通過了燕山大學倫理委員會的審批(批文編號:2019006)。
0 引言
視網膜退行性疾病使得視覺中樞無法正常接收到視覺信息,嚴重影響著患者的生活質量,增加了家庭和社會的經濟負擔[1]。它造成的視覺障礙經過標準的屈光矯正和藥物治療均無法有效改善其癥狀[2-3]。因此,如何開發治療視網膜退行性疾病的新技術成為了臨床的重要需求。目前常用的人工視網膜假體通常基于電神經刺激[4-6],是治療不同類型視網膜變性的先進技術。然而,這項技術需要將設備放入眼球,高度侵入性的手術不可避免地具有高風險和昂貴的成本。超聲刺激是一種有效且時空精確的激活視網膜的方法,超聲波刺激具有無創、高空間分辨率和高穿透深度優點[7-10],已被證明是一種有前途的無創神經刺激技術。先前研究使用超聲波調節視網膜神經節細胞(retinal ganglion cell,RGC)的放電活動,研究發現,超聲刺激離體小鼠視網膜能夠激活RGC,縮短了ON-RGC和OFF-RGC的光響應潛伏期,同時增強了部分RGC(ON-和OFF-亞型)的光響應并降低了其他RGC的光響應[11-12]。有研究使用離體的蠑螈視網膜來表征超聲波對完整神經回路的影響,發現超聲刺激可快速調節視覺敏感度。還有研究表明在正常視力或視網膜退化的失明大鼠中,對視網膜的直接超聲刺激可以激發上丘和初級視覺皮層(primary visual cortex,V1)[3]神經元活動。
以上研究表明超聲波可以激活健康和退化視網膜中的神經元。具體來說,超聲波可以引起局部神經元反應并在視覺回路中產生響應模式。但是目前關于超聲刺激視覺神經元如何調節視覺皮層的研究卻很少,超聲刺激視網膜時,視覺皮層如何進行信息編碼目前仍不清楚。
探究視覺系統的信息加工機制是神經科學領域的研究熱點[13-15]。人類的視覺系統主要由接收視覺信息的視網膜、傳遞視覺信息的外側膝狀體以及加工視覺信息的視覺皮層組成[16]。隨著神經成像技術的進步以及計算模型的發展,神經編碼和解碼逐漸成為研究視覺機制的主要手段[17-20]。視覺信息解碼旨在解決從皮層表征到視覺信息的逆向推理問題,基本方法為建立模式識別模型,根據記錄到的大腦響應推測出相應的視覺內容,也可以為腦機接口等前沿新興技術的應用提供基礎[19, 21]。因此我們十分有必要在超聲視覺刺激下,對視覺皮層的信息進行解碼。
超聲參數在超聲刺激中起著重要作用。先前的研究中,研究人員使用不同強度和刺激時間的超聲刺激蠕蟲(超聲參數:基頻10 MHz,頻率1 kHz,脈沖重復頻率,占空比50%),研究發現超聲刺激引起蠕蟲逆轉行為的成功率隨著超聲強度(0~1 MPa)的增加呈現近似線性的增加,且隨著刺激時間(50~400 ms)的增加而增加,但是超過閾值后會達到飽和[22]。另外,研究人員使用不同參數的超聲刺激小鼠的運動皮層激發肢體的運動響應,結果表明運動響應的成功率隨著聲強度(持續時間80 ms,頻率500 kHz,刺激強度0~16.8 W/cm2)和聲持續時間(強度4.2 W/cm2,頻率500 kHz,刺激時間0~320 ms)的增加而增加[23]。在我們先前的研究中發現超聲刺激小鼠運動皮層引起的局部場電位(local field potential,LFP)theta頻段的相對功率隨著超聲強度的增加而降低;低gamma頻段的相對功率隨著超聲強度的增加而增加,LFP的熵值隨著超聲強度和刺激時間的增加而減小(基頻500 kHz,脈沖重復頻率1 kHz,占空比40%,刺激強度0.2~1.1 W/cm2,刺激時間100~400 ms)[24]。我們的另一項研究還發現超聲刺激視覺皮層引起的神經元鈣活動的響應時間半高寬隨著刺激時間的增加而增加(刺激強度0.45 MPa,基頻500 kHz,脈沖重復頻率1 kHz,占空比10%,刺激時間300~700 ms)[25]。以上研究表明超聲刺激對神經的調控效果與超聲參數密切相關。但是超聲視覺刺激引起的視覺皮層神經活動是否與超聲參數(強度和刺激時間)具有依賴關系尚不清楚。
為了回答以上兩個問題,我們使用不同強度和刺激時間的超聲刺激小鼠的視網膜,首次探討了超聲視網膜刺激對視覺皮層的調節作用與超聲參數之間的定量關系。它能夠幫助理解超聲刺激對視覺系統功能的調控機制。本研究旨在明確參數選擇規律,以期推動個體化多參數調控治療視覺神經退化、視網膜病變等疾病研究方向的開展。
1 材料和方法
1.1 動物和分組
本實驗中,使用了20只C57BL/6小鼠(均為雄性,體重20~25 g,北京維通利華實驗動物科技有限公司)。所有實驗環節均符合燕山大學動物倫理要求。在實驗開始前7天起至實驗結束期間,均將小鼠飼養在12小時光照/黑暗循環的標準籠具中,按需提供鼠糧和飲用水。并將小鼠隨機分為超聲刺激強度組和超聲刺激時間組,每組10只。對小鼠視網膜進行超聲刺激,記錄V1的LFP。
1.2 動物實驗流程
利用移動式小動物麻醉機(R530,深圳市瑞沃德生命科技有限公司)對小鼠進行麻醉,異氟烷流通速度設置為2%,氧氣流通速度設置為0.5 L/min。利用小鼠適配器(
1.3 超聲刺激系統
超聲刺激系統由任意波形函數發生器1(DG1022U,普源精電科技股份有限公司)、任意波形函數發生器2(AFG
圖1
超聲刺激系統、數據采集系統及超聲刺激參數
a. 超聲刺激系統和數據采集系統;b. 超聲刺激參數
Figure1. The ultrasound stimulation system, data acquisition system, and ultrasound stimulation parametersa. ultrasound stimulation system and data acquisition system; b. ultrasound stimulation parameters
1.4 數據采集系統
使用多通道數據采集系統(Apollo,Bio-Signal Technologies,美國)記錄超聲刺激位點和腦電信號。如圖1所示,從小鼠V1采集到的信號經由前置放大器放大,通過采集系統處理得到腦電信號。采集過程中,LFP的采樣率設置為1 kHz。
1.5 LFP預處理
對于鎳鉻電極采集到的16通道LFP信號,去除干擾較大通道的LFP信號,對余下LFP信號進行帶通濾波處理,帶通濾波范圍設置為0.5~200 Hz,并濾掉3 Hz的運動偽跡干擾。
1.6 時頻圖和功率分析
在時頻圖分析過程中采用Welch算法[26],結合刺激位點獲取超聲刺激–1~2 s的LFP數據,分別進行時頻圖處理,對于得到的多通道時頻圖結果進行平均,截取0.5~45 Hz頻段的時頻圖。在功率分析過程中,分別計算了LFP的delta、theta、alpha、beta、低gamma頻段在超聲刺激–1~0、0~1、1~2 s的功率,并觀察不同刺激強度/時間下,各個頻段的腦電功率隨強度/時間變化的情況。
1.7 樣本熵分析
我們參考了先前的文獻[27],分別計算了LFP的delta、theta、alpha、beta、低gamma頻段在超聲刺激–1~0、0~1、1~2 s的樣本熵,并觀察不同刺激強度/時間下,各個頻段的樣本熵隨強度/時間參數變化的情況。
1.8 統計分析
采用多因素方差分析對數據進行統計學分析。檢驗水準為0.05。
2 結果
2.1 不同強度的超聲刺激視網膜時V1的LFP
使用不同強度的超聲刺激視網膜,對采集到的V1的LFP進行分析。圖2a中依次呈現了不同刺激強度下(0.1、0.2、0.3、0.4、0.5 MPa)的LFP及時頻圖,可以發現,隨著超聲刺激強度的增加,LFP的幅值呈現上升趨勢,并且在時頻圖的0~1 s,可以觀察到隨著超聲刺激強度的增加,delta頻段的功率有明顯的增強,theta頻段略微增強。如圖2b所示,我們計算了LFP的峰峰值(最高正峰值和最低負峰值的差值),隨著超聲刺激強度的增加,峰峰值呈現明顯的上升趨勢(R2 = 0.694 7)。如圖2c所示,隨著超聲刺激強度的增加,delta頻段的功率同其他頻段相比變化最為明顯,呈現明顯的上升趨勢(R2=0.520 4)。如圖2d所示,隨著超聲刺激強度的增加,theta頻段的功率呈現上升趨勢(R2 =
圖2
不同強度超聲刺激視網膜時V1的LFP結果
a. LFP和時頻圖;b. 峰峰值;c-g. delta、theta、alpha、beta、low gamma的功率(mean ± sem,
a. the LFP and time-frequency diagram; b. the peak to peak amplitude; c-g. the power of delta, theta, alpha, beta, and low gamma (mean ± sem,
2.2 不同強度的超聲刺激視網膜時V1的樣本熵
使用不同強度的超聲刺激視網膜時,我們還分析了不同頻段LFP的樣本熵。如圖3a~b所示,隨著超聲刺激強度的增加,delta、theta頻段的樣本熵均呈現下降趨勢(delta:R2 = 0.155 3;theta:R2 = 0.191 3)。如圖3c~e所示,隨著超聲刺激強度的增加,alpha、beta、低gamma頻段的樣本熵無明顯變化。以上結果表明超聲刺激視網膜在V1誘發的LFP低頻段的非線性特征隨著超聲強度的增加而降低。
圖3
不同強度超聲刺激視網膜時V1的樣本熵結果
a-e. delta、theta、alpha、beta、low gamma頻段的樣本熵(mean ± sem,
a-e. the sample entropy of delta, theta, alpha, beta, and low gamma (mean ± sem,
2.3 不同時間的超聲刺激視網膜時V1的LFP
使用不同時間的超聲刺激視網膜,對采集到的V1的LFP進行分析。圖4a中依次呈現了不同刺激時間下(0.1、0.2、0.3、0.4、0.5 s)的LFP及時頻圖,可以發現,隨著超聲刺激時間的增加,LFP的幅值也呈現上升趨勢,并且在時頻圖的0~1 s,觀察到delta頻段的功率有明顯增強。如圖4b所示,我們計算了LFP的峰峰值,隨著超聲刺激時間的增加,峰峰值呈現上升趨勢,與不同強度超聲刺激時比較,上升較為緩慢,且隨著超聲刺激時間進一步增加,峰峰值上升趨勢減弱。如圖4c所示,隨著超聲刺激時間的增加,delta頻段的功率整體呈現上升趨勢,刺激時間為0.1~0.3 s時,上升趨勢明顯,0.3 s后上升趨勢開始減弱,0.4~0.5 s之間變化不明顯。如圖4d~g所示,隨著超聲刺激時間的增加,theta、alpha、beta、低gamma頻段的功率無明顯變化。以上結果同樣表明超聲刺激視網膜能夠誘發V1的LFP變化,即引起了神經活動;而且,超聲刺激視網膜誘發的低頻神經活動隨著超聲刺激時間的增加而增加,但是在時間增加到一定程度后一些特征的增加趨勢減弱。
圖4
不同時間超聲刺激視網膜時V1的LFP結果
a. LFP和時頻圖;b. 峰峰值;c-g. delta、theta、alpha、beta、low gamma的功率(mean ± sem,
a. the LFP and time-frequency diagram; b. the peak to peak amplitude; c-g. the power of delta, theta, alpha, beta, and low gamma (mean ± sem,
2.4 不同時間超聲刺激視網膜時V1的樣本熵
使用不同時間的超聲刺激視網膜時,我們還分析了V1腦區不同頻段LFP的樣本熵。如圖5a所示,隨著超聲刺激時間的增加,delta頻段的樣本熵整體呈現上升趨勢(R2=0.070 3)。如圖5b所示,隨著超聲刺激時間的增加,theta頻段的樣本熵呈現下降趨勢,且隨著超聲刺激時間的進一步增加,下降趨勢減弱。如圖5c~e所示,隨著超聲刺激時間的增加,alpha、beta、低gamma頻段的樣本熵無明顯變化。以上結果表明超聲刺激視網膜可在V1誘發LFP低頻段的非線性特征的變化。該特征在delta頻段隨著超聲刺激時間的增加而升高,在theta頻段隨著超聲刺激時間的增加而降低。
圖5
不同時間超聲刺激視網膜時V1的樣本熵結果
a-e. delta、theta、alpha、beta、low gamma頻段的樣本熵(mean ± sem,
a-e. the sample entropy of delta, theta, alpha, beta, and low gamma (mean ± sem,
3 討論
本文中,我們對小鼠視網膜進行超聲刺激,采集V1的LFP,并對于超聲刺激前后(–1~2 s)的時頻圖、功率及樣本熵進行處理和分析。結果表明,利用不同強度、不同時間的超聲刺激小鼠視網膜時,V1的神經活動會發生改變。隨著超聲強度的增大,V1的LFP峰峰值變大,低頻段的功率增強,非線性特征減弱;隨著超聲刺激時間的增加,V1的LFP峰峰值變大,delta頻段的功率增強但刺激時間增加到一定程度后增強趨勢減弱,delta頻段非線性特征增強,theta頻段非線性特征減弱。以上結果都表明在刺激視網膜時,超聲強度、超聲時間參數的改變都會對V1的神經活動產生調節作用。
我們發現,對于峰峰值、delta頻段功率的變化,超聲刺激強度組的變化趨勢強于超聲刺激時間組。但是何種超聲強度和刺激時間組合對于患者視覺系統的調節作用是最佳的,目前尚不明確,需要結合臨床進一步探討。通過其他學者的研究,我們也注意到如何最優利用超聲刺激,并不局限于超聲參數這一個因素。先前的研究表明,多個視覺區域在功能和視覺感知上是相互聯系的,比如刺激V2或者V3也可以影響V1[28]。視神經、視覺丘腦以及視覺皮層也可以作為研究視覺系統的靶點,已有學者通過視神經內刺激來為全盲患者解決視覺問題[29-30]。在今后的研究中,結合不同的刺激靶點研究不同超聲參數對于視覺系統的調節作用,是一個有價值的研究方向,可為臨床視網膜退化性疾病、視覺損傷疾病的治療提供一定的參考。
先前的研究表明眼部活動和腦部活動之間聯系密切[31],且視網膜神經元可表達諸如大鉀離子(big potassium,BK)、瞬時受體電位(transient receptor potential,TRP)等在內的多個機械敏感離子通道[32]。我們推測低強度超聲刺激可能通過調節視網膜的機械敏感離子通道改變了不同類型神經元之間的分布模式,通過RGC的活動,進而影響神經信號向大腦視覺皮層的傳遞,最終改變視覺皮層的神經活動模式。我們的研究表明,改變超聲刺激強度和時間可以實現對于V1神經活動不同程度的調節。在超聲刺激視網膜改善視覺功能的工作中,希望本研究可以推動多參數協同治療的發展,在未來的研究中可以進一步探索超聲在視網膜退行疾病中的應用潛力,并深入探索視網膜-初級視覺皮層通路可塑性。
重要聲明
利益沖突聲明:本文全體作者均聲明不存在利益沖突。
作者貢獻聲明:王倩倩負責實驗設計、數據采集、數據處理和論文初稿,袁毅負責實驗監督以及論文審查和編輯,閆佳慶負責實驗監督和數據處理。
倫理聲明:本研究通過了燕山大學倫理委員會的審批(批文編號:2019006)。