引用本文: 楊春生, 陳禰, 武恒敏, 張鐵成, 王寶蘭. 慢性晝夜節律紊亂對大鼠膝骨關節炎樣軟骨損傷的機制研究. 華西醫學, 2024, 39(6): 891-898. doi: 10.7507/1002-0179.202403266 復制
版權信息: ?四川大學華西醫院華西期刊社《華西醫學》版權所有,未經授權不得轉載、改編
膝骨關節炎(knee osteoarthritis, KOA)是一種臨床上最常見的骨關節疾病。流行病學研究發現,長期輪班工作與髖部、腕部骨折、KOA 的風險之間存在相關性,這表明慢性晝夜節律被破壞可能會增加關節軟骨退化的易感性[1-2]。晝夜節律是一種適應性機制,由生物鐘主導調控。基本螺旋體型蛋白 1(basic helix-loop-helix ARNT like 1, BMAL1)作為生物鐘網絡的核心,是多種生理功能的重要調節劑,包括新陳代謝、睡眠、體溫、血壓和軟骨發育等。研究發現,慢性晝夜節律紊亂可引起膝關節軟骨中 Bmal1 表達下調,下游多條信號通路發生變化,是 KOA 的一個危險因素[3-4]。在軟骨特異性 Bmal1 敲除小鼠生長板中軟骨細胞增殖和存活率明顯降低,凋亡軟骨細胞的數量較高[5]。而細胞凋亡和異常分化主要受細胞周期調控,細胞周期本身也具有節律性,與生物鐘之間存在耦合關系。生物鐘基因可直接或間接控制肝臟、泌尿生殖等多個器官組織中細胞周期基因和蛋白的表達,如 WEE1 細胞周期 G2 檢測點激酶(WEE1 G2 checkpoint kinase, Wee1)、細胞周期蛋白 E 和細胞周期蛋白 B1(cyclin B1, CCNB1)等[6-8]。然而,膝關節軟骨中生物鐘和細胞周期相互影響的機制尚不完全清楚。因此,了解晝夜節律振蕩和細胞周期之間的關系,對于 KOA 的機制研究具有重要意義。
1 材料與方法
1.1 材料
1.1.1 實驗動物
40 只 5~6 周齡無特定病原體的雄性 SD 大鼠,體重 160~210 g,購自新疆醫科大學動物中心。本研究所有動物的實驗設計均獲得新疆醫科大學第一附屬醫院動物倫理委員會批準,倫理審批號:A2310-22。
1.1.2 主要試劑與儀器
番紅固綠染色試劑盒(北京索萊寶科技有限公司);聚合酶鏈反應(polymerase chain reaction, PCR)試劑盒(北京全式金生物技術有限公司);一抗[BMAL1、WEE1、細胞周期依賴性激酶 1(cyclin-dependent kinase 1, CDK1)、CCNB1、細胞凋亡調節劑 2(apoptosis regulator 2, BCL2)、BCL2 相關 X 蛋白(BCL2-associated X protein, BAX),中國武漢三鷹生物技術有限公司];二抗(山羊抗兔)、蛋白裂解液、蛋白濃度檢測試劑盒、蘇木素液和細胞凋亡檢測試劑盒(武漢塞維爾生物科技有限公司);SEDI PCR 儀(英國威泰克公司);PowerPac HC 蛋白電泳儀(美國 BIO-RAD 公司)。
1.2 方法
1.2.1 實驗分組和晝夜節律紊亂模型建立
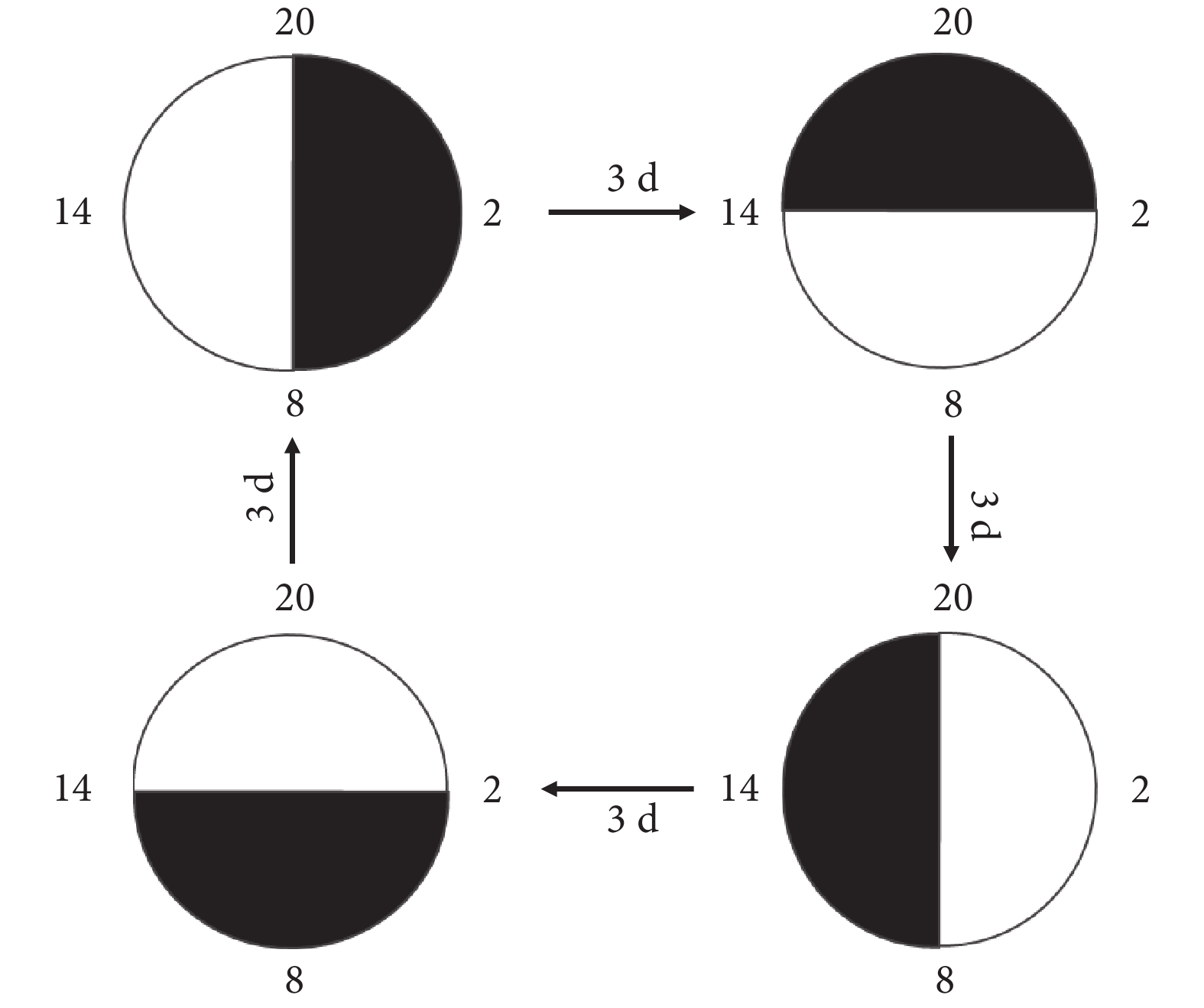
40 只大鼠隨機分為晝夜節律正常組(正常組)、晝夜節律紊亂組(紊亂組)、Bmal1 過表達慢病毒感染的晝夜節律紊亂組(Bmal1 上調組)和 Bmal1 過表達慢病毒陰性感染的晝夜節律紊亂組(Bmal1 陰性感染組),每組 10 只。實驗期間,4 組大鼠明暗周期均為 12 h,起始 8 點開燈,20 點關燈。正常組始終保持 8 點開燈和 20 點關燈,紊亂組、Bmal1 上調組和 Bmal1 陰性感染組每隔 3 d 提前 6 h 開燈,此為 1 個循環,共進行 5 個循環,持續約 60 d,大鼠在不斷的明暗周期中發生慢性晝夜節律紊亂,見圖1。
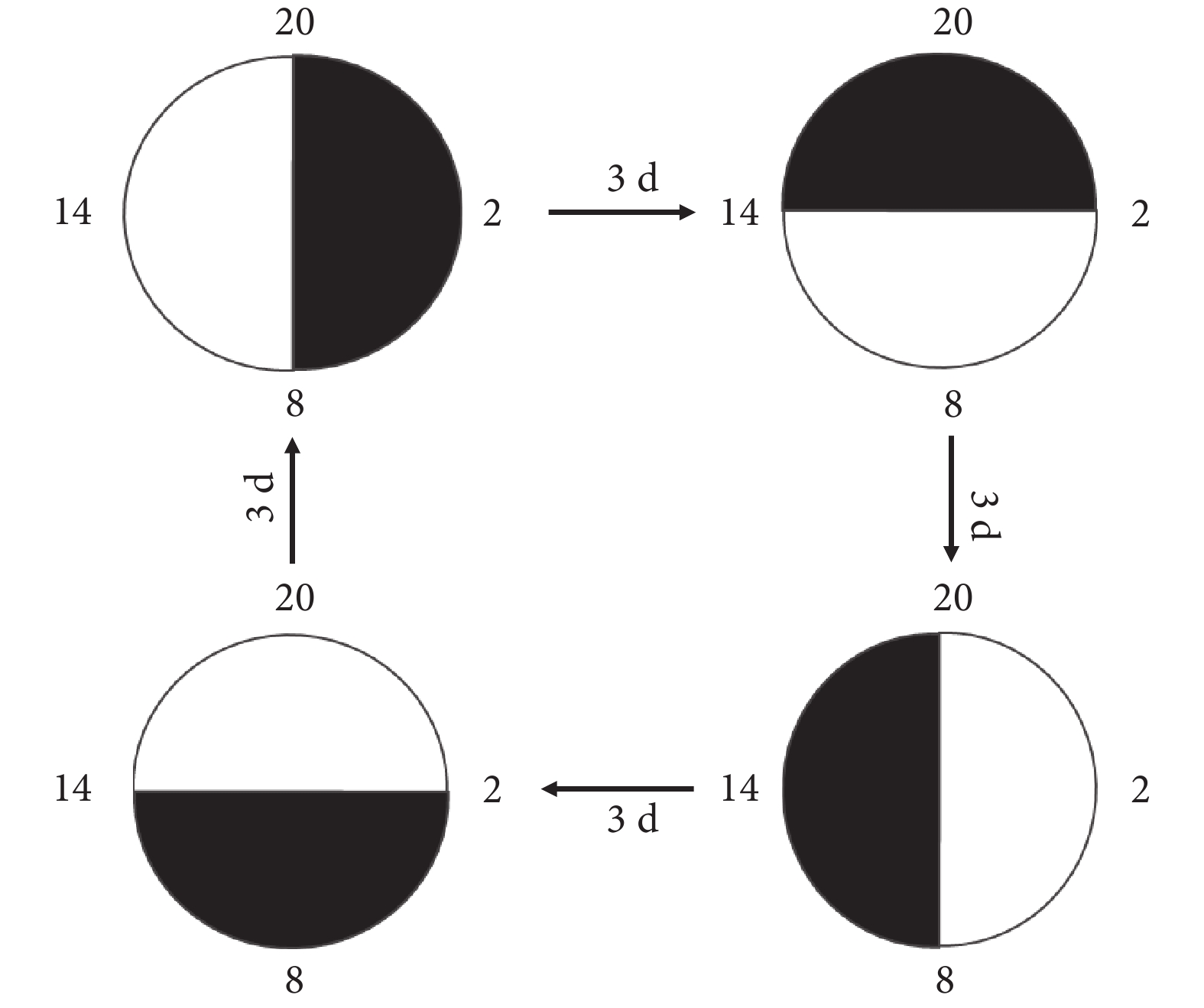 圖1
晝夜節律紊亂模型
圖1
晝夜節律紊亂模型
1.2.2 Bmal1 過表達和陰性慢病毒關節腔注射模型建立
首先制備 Bmal1 過表達和陰性慢病毒的稀釋液,滴度均為 1×108 TU/mL,置于–80°C 冰箱保存備用。晝夜節律紊亂模型開始建立的第 1 天,將 Bmal1 上調組和 Bmal1 陰性感染組的大鼠麻醉后,在左后膝關節腔內分別注射 Bmal1 過表達慢病毒和 Bmal1 過表達陰性慢病毒各 50 μL,30 d 后重復注射 1 次。
1.2.3 番紅固綠染色
模型建立后大鼠深度麻醉后放血致死,無菌條件下手術摘取膝關節,置于組織固定液中,常規脫水、透明、包埋、切片后,進行番紅固綠染色,顯微鏡下觀察軟骨病理變化。
1.2.4 原位末端轉移酶標記法(TdT-mediated dUTP nick-end labeling, TUNEL)染色
石蠟切片脫蠟至水,蛋白酶工作液覆蓋組織、37℃溫箱孵育,切片置于磷酸鹽緩沖液(phosphate buffered saline, PBS)洗滌、滴加破膜液覆蓋組織、常溫孵育,切片置于 PBS 中洗滌、滴加 buffer 覆蓋組織、常溫孵育。混合好的工作液加到組織上,于濕盒內 37℃恒溫箱孵育、PBS 洗滌、滴加 4’, 6-二脒基-2-苯基吲哚染液,避光室溫孵、PBS 中洗滌。切片甩干、抗熒光淬滅封片劑封片。熒光顯微鏡下觀察并采集圖像。
1.2.5 膝關節軟骨組織的免疫組織化學檢測
軟骨組織經多聚甲醛固定后石蠟包埋切片,脫蠟、脫水、洗片后,過氧化氫液去除內源性過氧化物酶,檸檬酸鈉和乙二胺四乙酸緩沖液進行抗原修復,一抗(BMAL1、WEE1、CDK1、CCNB1、BAX 和 BCL2)孵育過夜。第 2 天,二抗常溫孵育,二氨基聯苯胺顯色、稀鹽酸分色、蘇木素反藍,最后行脫水、透明、封片、采圖。所有染色切片通過 Image J 軟件測量陽性細胞面積進行差異性分析。
1.2.6 膝關節軟骨組織的 mRNA 表達檢測和相關性分析
主要應用 PCR 技術,將軟骨組織在加入液氮的研缽內研磨成粉,應用軟骨 RNA 柱式提取試劑盒,進行 Bmal1、Wee1、Bcl2、Cdk1、Ccnb1、Bax、白細胞介素 1(interleukin 1, Il1)、白細胞介素 6(interleukin 6, Il6)、腫瘤壞死因子(tumor necrosis factor, Tnf)和基質金屬蛋白酶 13(matrix metallopeptidase 13, Mmp13)的 RNA 提取,逆轉錄為互補 DNA 后行基因擴增。PCR 采用兩步法,擴增條件:94℃預變性 30 s,94℃變性 5 s,60℃退火和延伸 30 s,共 40 個循環。根據檢測結果的循環閾值,計算 mRNA 的相對表達量,進行差異性分析。根據實驗組中 mRNA 的相對表達量,使用 OriginPro 2021 軟件的 Correlation Plot 插件進行相關性分析。
1.2.7 膝關節軟骨組織的蛋白質印跡法檢測
將研磨后軟骨加入放射免疫沉淀測定法裂解液和蛋白磷酸酶抑制劑,在冰上提取細胞總蛋白,BCA 試劑盒測定蛋白濃度。每個蛋白樣品等量在 10% 聚丙烯酰胺凝膠上樣電泳分離,進行轉膜,5% 脫脂奶粉封閉后添加一抗(BMAL1、WEE1、CDK1、CCNB1、BAX 和 BCL2),在 4℃下過夜。PBS 溶液加上 Tween-20(PBS with Tween-20, PBST)洗滌 3 次后孵育二抗,在室溫下避光處理 1 h。PBST 洗膜 3 次,5 min/次,并用 PBS 沖洗以去除成像前的 Tween-20,化學發光成像。用 Image J 軟件進行灰度值分析。
1.3 統計學方法
應用 SPSS 23.0 軟件進行實驗數據的處理分析,統計數據服從正態分布,計量資料以均數±標準差表示,多組間比較應用單因素方差分析,多重比較采用 Tukey 法。應用 GraphPad Prism 9.0 軟件作圖,OriginPro 2021 軟件進行 Pearson 相關性分析。雙側檢驗水準 α=0.05。
2 結果
2.1 番紅固綠染色
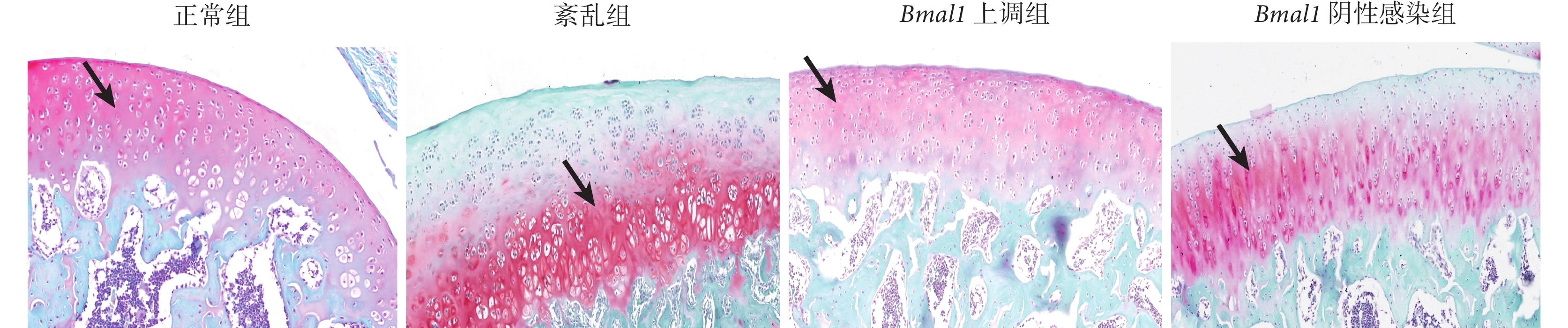
正常組軟骨基質厚度正常,呈均勻的紅色,軟骨下骨為綠色。紊亂組軟骨基質破壞,蛋白聚糖丟失明顯,呈不均勻紅色;Bmal1 上調組軟骨基質厚度基本正常,蛋白聚糖無明顯丟失,紅色略欠均勻,軟骨下骨為綠色;Bmal1 陰性感染組軟骨出現基質破壞和蛋白聚糖丟失,呈不均質紅色。見圖2。
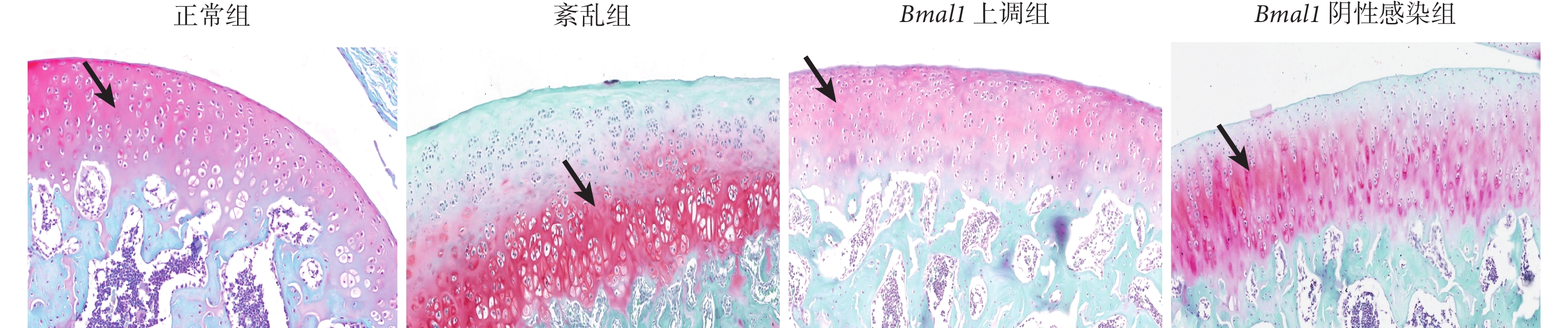 圖2
各組大鼠關節軟骨番紅固綠染色病理圖片(×100)
圖2
各組大鼠關節軟骨番紅固綠染色病理圖片(×100)
黑箭指向為蛋白聚糖
2.2 TUNEL 染色結果
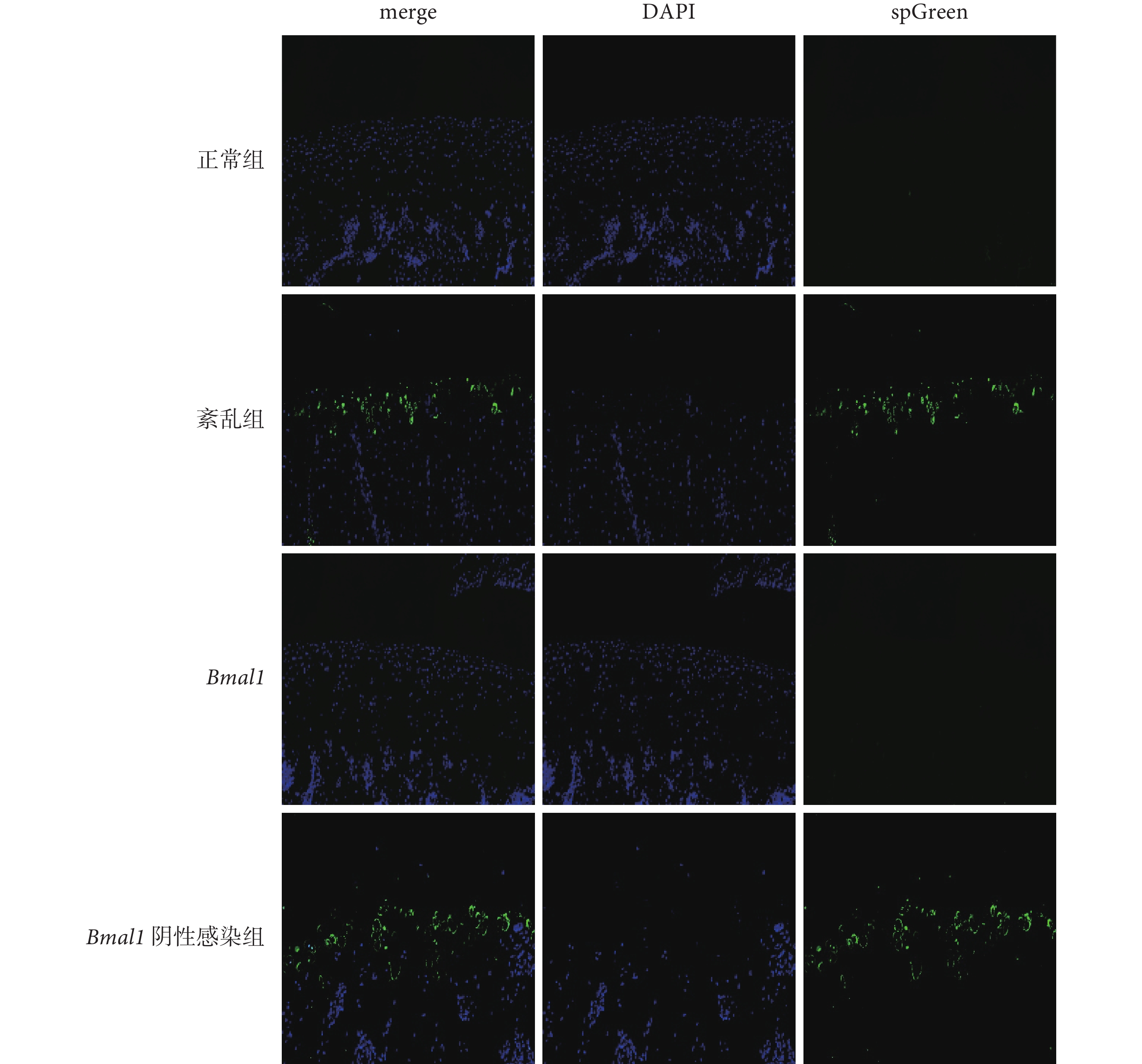
凋亡的陽性細胞被染為綠色,形態、大小各異,散在不均勻地分布于正常細胞之間。與正常組和 Bmal1 上調組相比,紊亂組和 Bmal1 陰性感染組凋亡陽性細胞比率明顯升高,差異有統計學意義(P<0.001);正常組和 Bmal1 上調組相比,以及紊亂組和 Bmal1 陰性感染組相比,凋亡陽性細胞比率差異均無統計學意義(P>0.05)。見圖3 和表1。
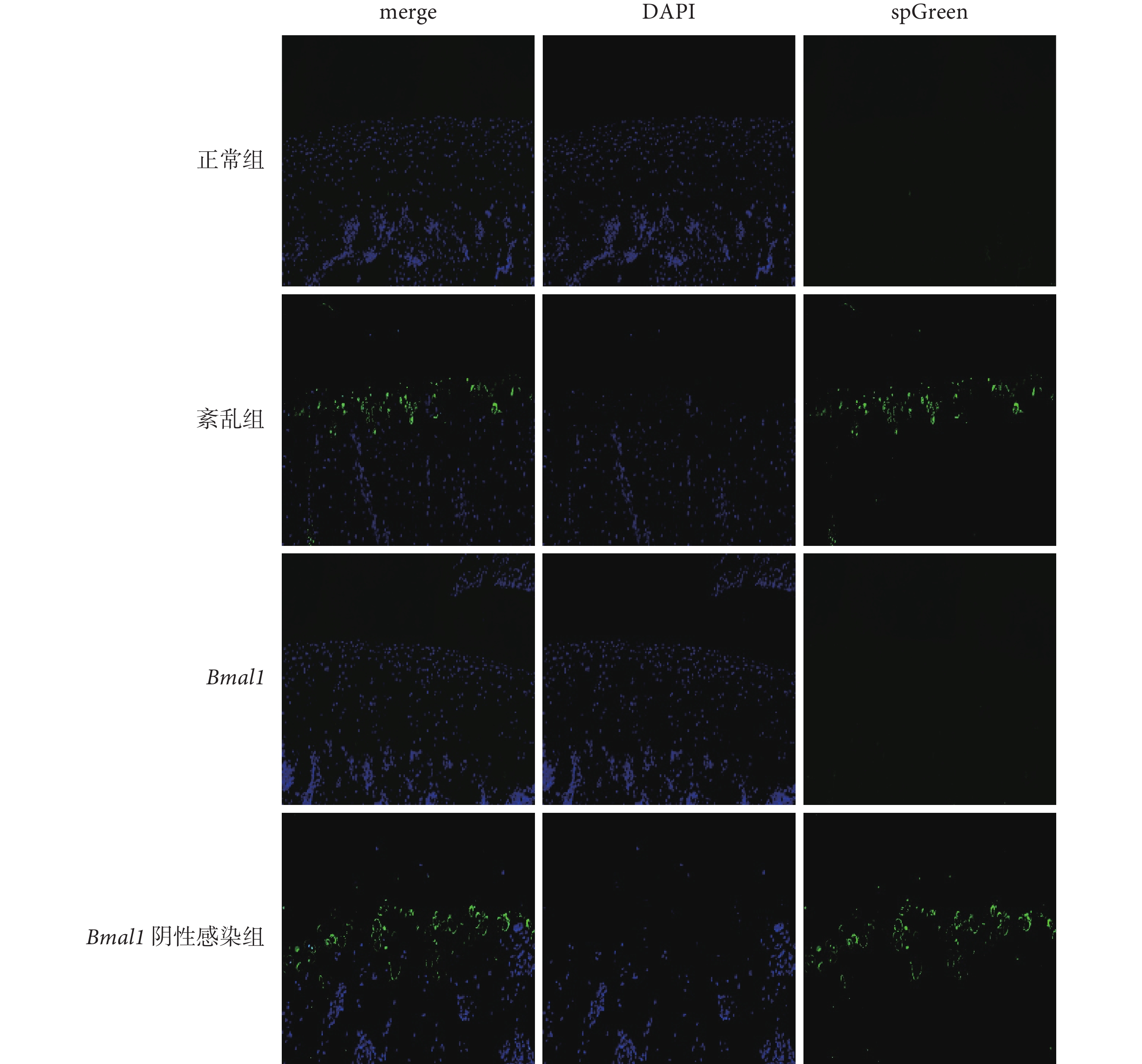 圖3
各組大鼠關節軟骨 TUNEL 染色結果(×200)
圖3
各組大鼠關節軟骨 TUNEL 染色結果(×200)
藍色為正常軟骨細胞,綠色為凋亡的軟骨細胞。TUNEL:原位末端轉移酶標記法;merge:合并;DAPI:4’, 6-二脒基-2-苯基吲哚;spGreen:新型無毒核酸染料
2.3 各組軟骨組織中免疫組織化學分析結果
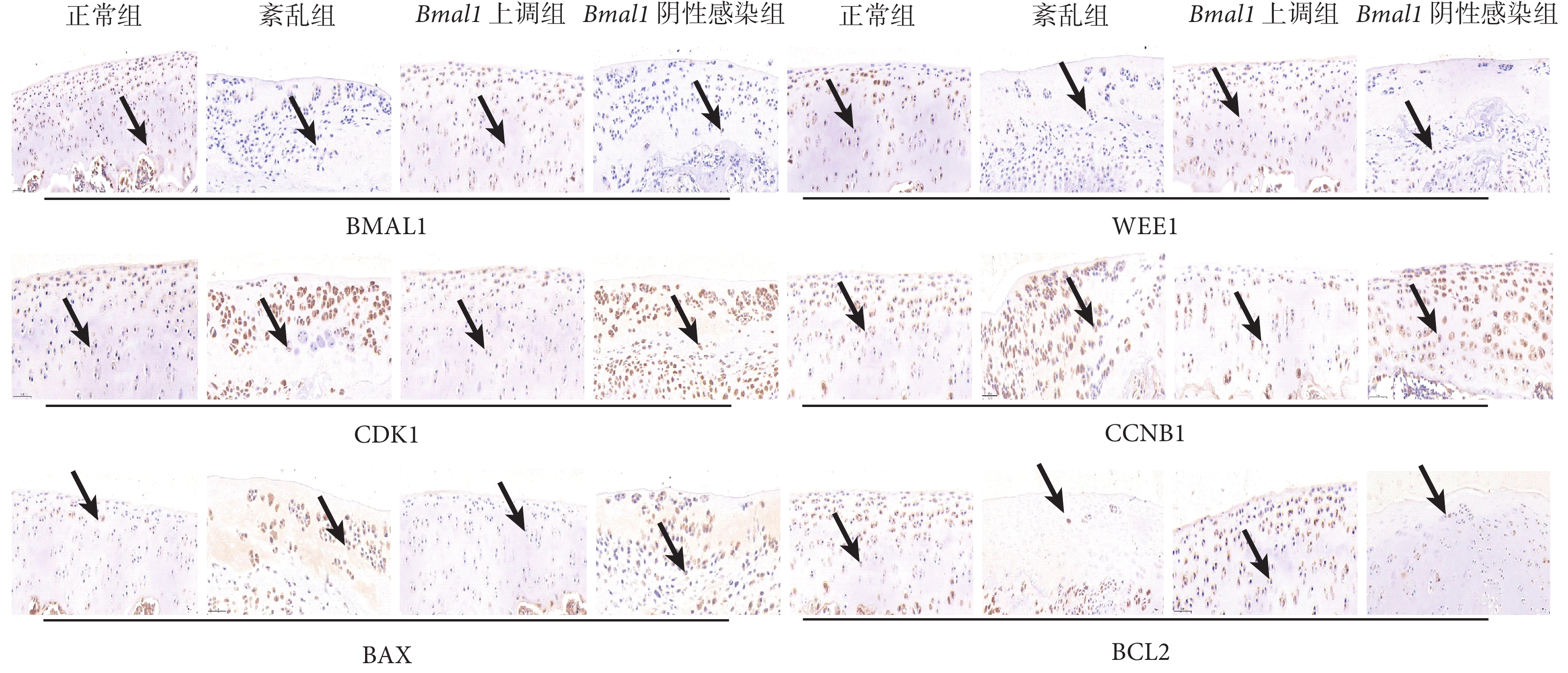
與正常組和 Bmal1 上調組相比,紊亂組和 Bmal1 陰性感染組關節軟骨中 BMAL1、WEE1 和 BCL2 蛋白的相對表達水平下降,CDK1、CCNB1 和 BAX 的蛋白相對表達水平增加,差異有統計學意義(P<0.05);正常組和 Bmal1 上調組相比,以及紊亂組和 Bmal1 陰性感染組相比,上述蛋白的相對表達水平差異均無統計學意義(P>0.05)。見表1、圖4。
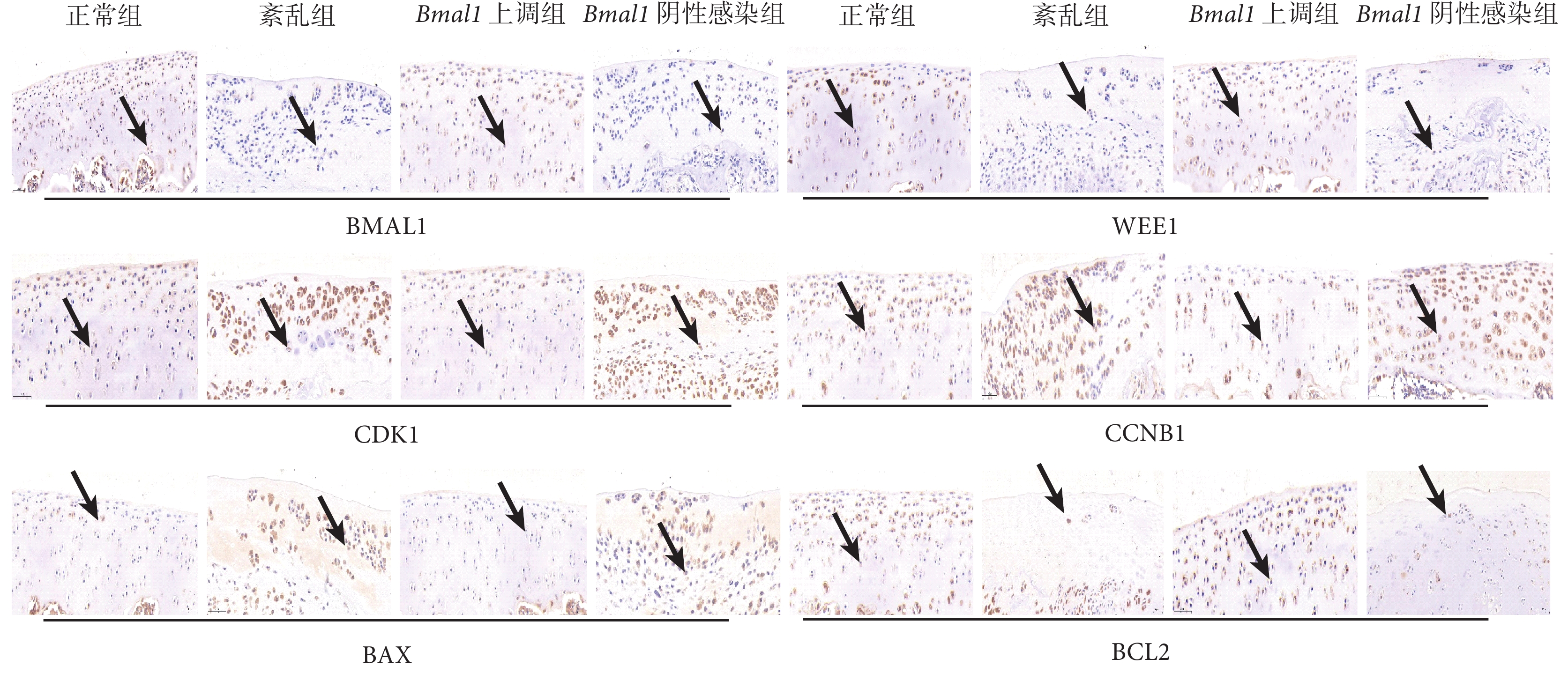 圖4
細胞周期相關蛋白免疫組織化學結果(×200)
圖4
細胞周期相關蛋白免疫組織化學結果(×200)
黑箭指向為染色陽性軟骨細胞
2.4 各組軟骨組織的逆轉錄 PCR 檢測和相關性分析
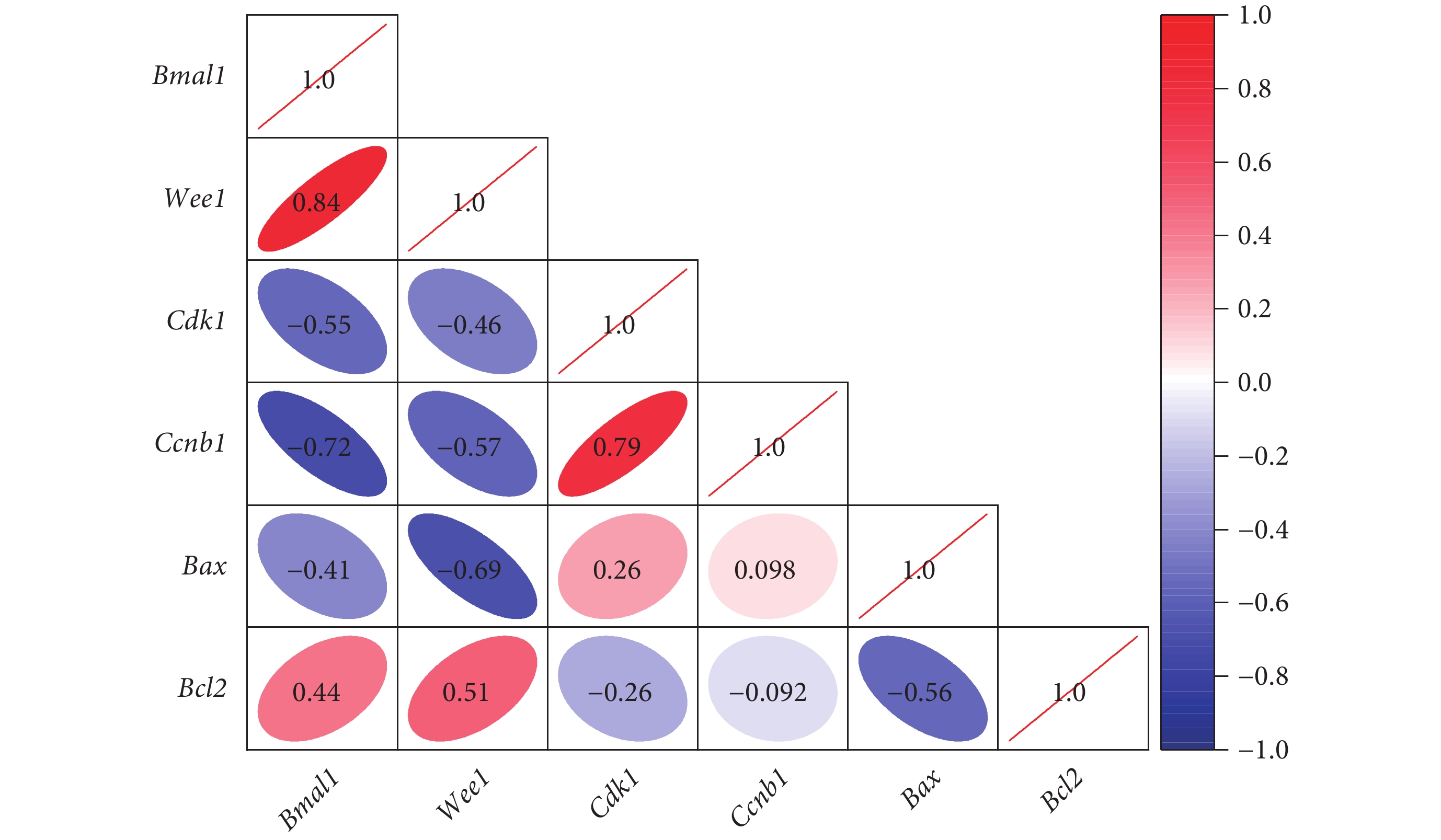
與正常組和 Bmal1 上調組相比,紊亂組和 Bmal1 陰性感染組關節軟骨中 Bmal1、Wee1 和 Bcl2 的 mRNA 相對表達量下降,Cdk1、Ccnb1、Bax、Il1、Il6、Tnf 和 Mmp13 的 mRNA 相對表達量增加,差異有統計學意義(P<0.05);正常組和 Bmal1 上調組相比,以及紊亂組和 Bmal1 陰性感染組相比,上述基因的 mRNA 相對表達量差異均無統計學意義(P>0.05),見表2。相關性分析顯示,Bmal1 與 Wee1、Bcl2 呈正相關,相關性系數分別為 0.84 和 0.44(P<0.01);與 Cdk1、Ccnb1、Bax 呈負相關,相關性系數分別為?0.55、?0.72、?0.41(P<0.01),見圖5。
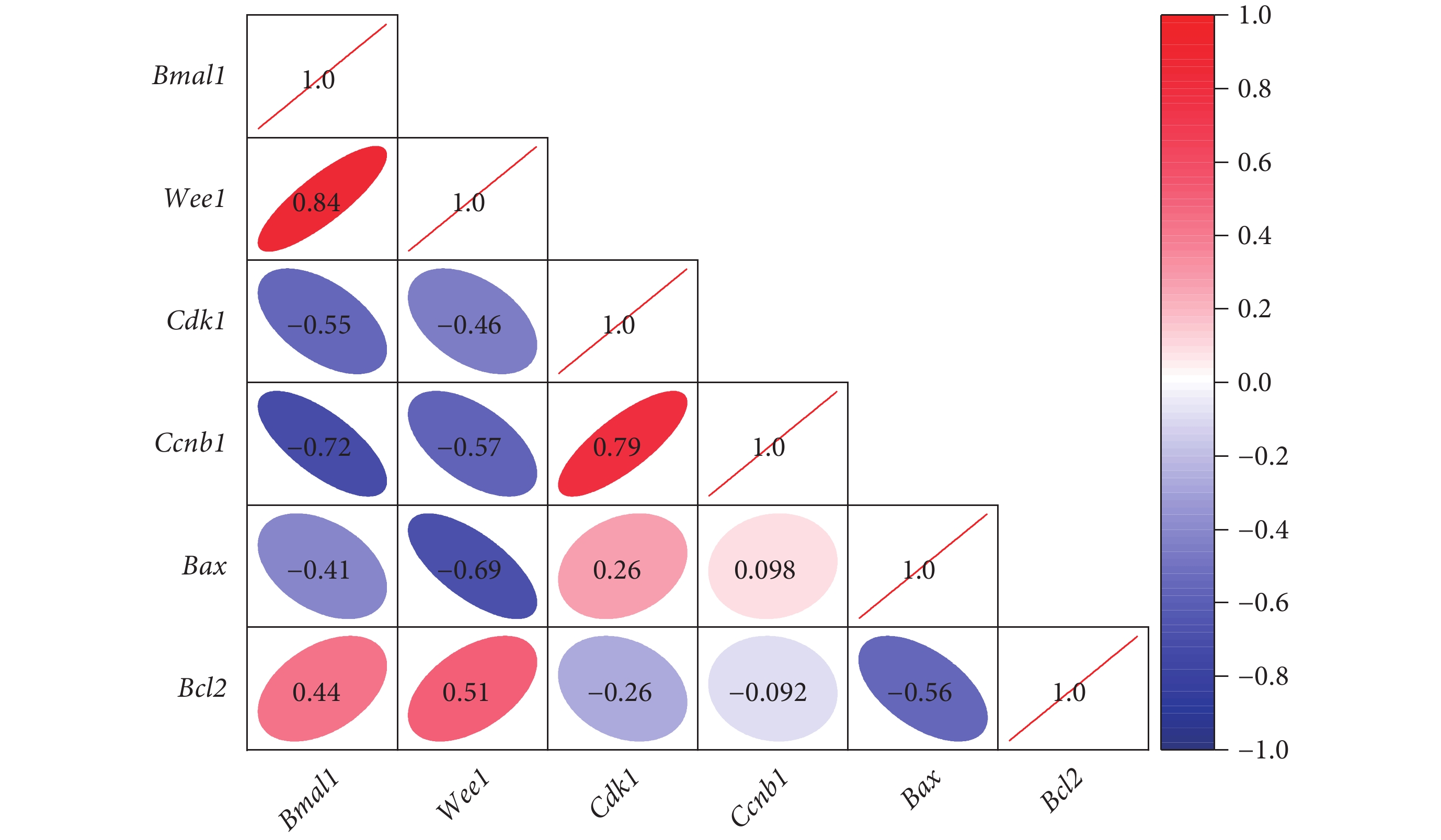 圖5
細胞周期相關基因相關性分析
圖5
細胞周期相關基因相關性分析
2.5 各組軟骨組織蛋白質印跡分析結果
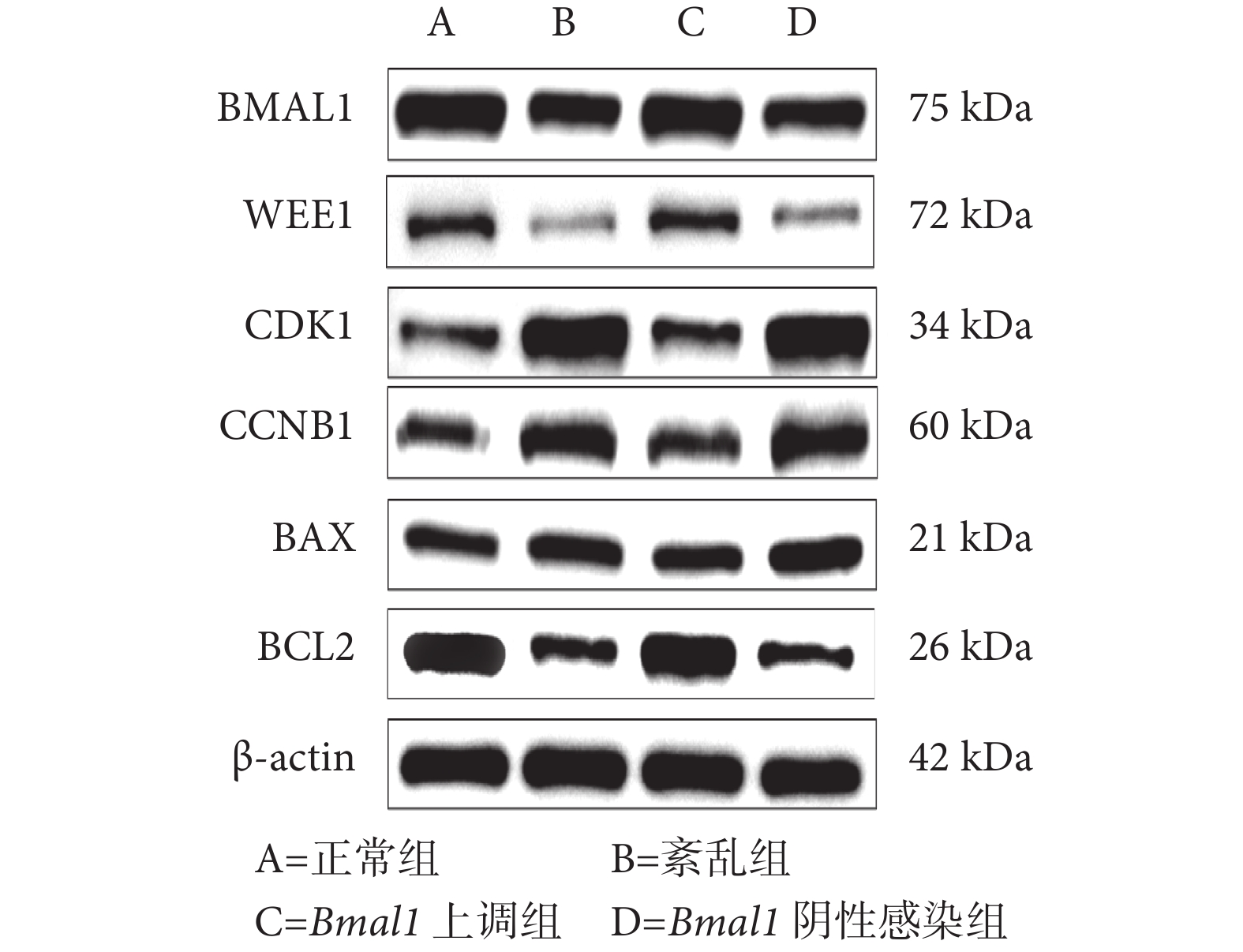
與正常組和 Bmal1 上調組相比,紊亂組和 Bmal1 陰性感染組中 BMAL1、WEE1 和 BCL2 蛋白的相對表達水平下降,CDK1、CCNB1 和 BAX 的蛋白相對表達水平增加,差異有統計學意義(P<0.05);正常組和 Bmal1 上調組相比,以及紊亂組和 Bmal1 陰性感染組相比,上述蛋白的相對表達水平差異均無統計學意義(P>0.05),見表3 和圖6。
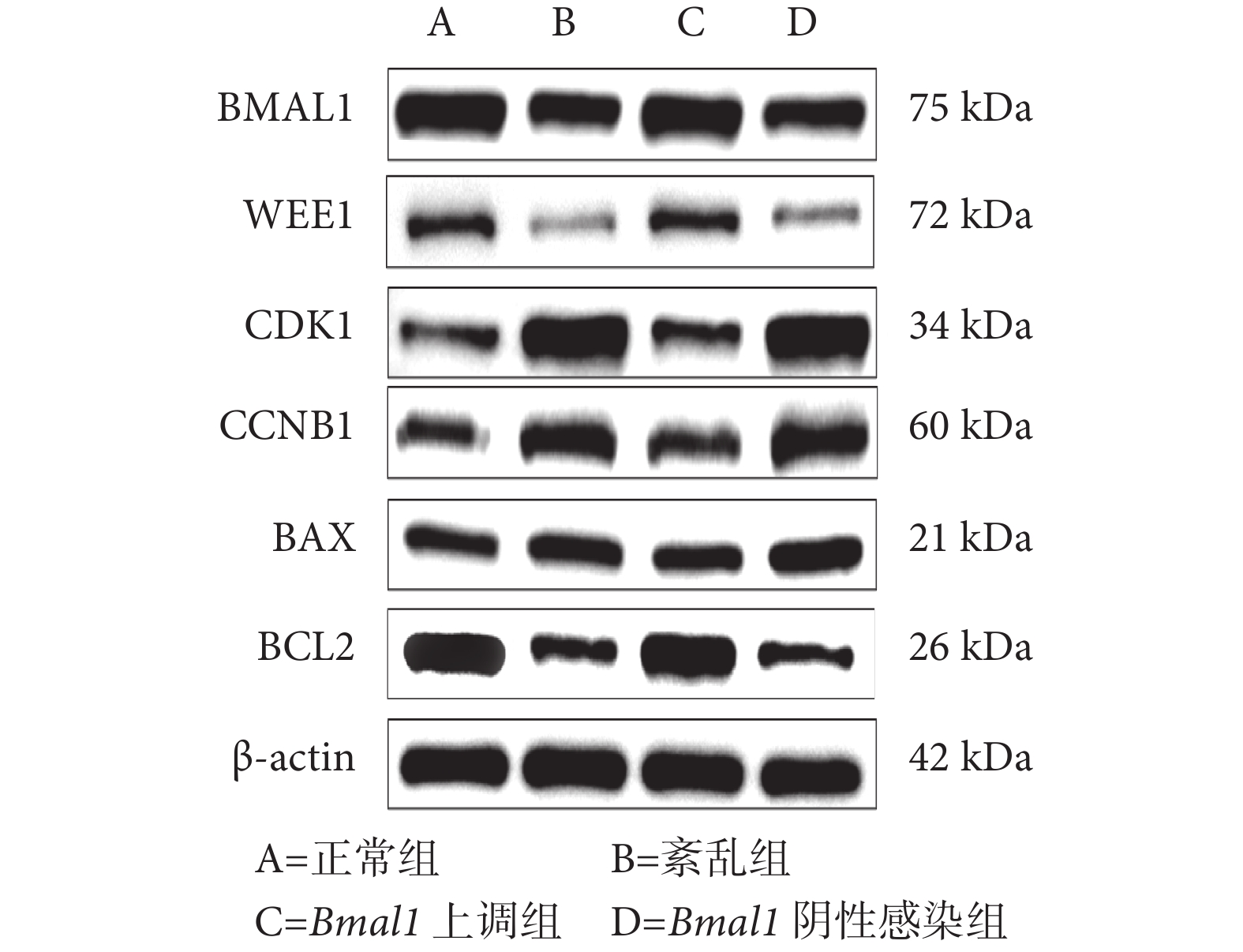 圖6
蛋白質印跡檢測
圖6
蛋白質印跡檢測
3 討論
本研究發現,當慢性晝夜節律紊亂引起 Bmal1 下調,給予 Bmal1 上調慢病毒干預后,關節軟骨中 Bmal1、Wee1 和 Bcl2 的表達上調,Cdk1、Ccnb1、Bax、Il1、Il6、Tnfa 和 Mmp13 的表達下調,蛋白聚糖丟失改善,軟骨細胞凋亡比率下降。我們也據此推測,Bmal1、Wee1 和 Bcl2 的升高可能會阻滯骨關節炎的進程,Cdk1、Ccnb1、Bax 的升高會加劇骨關節炎的發展。慢性晝夜節律紊亂后,生物鐘基因 Bmall 可能通過影響細胞周期相關基因,在軟骨細胞的生長、增殖、分裂和凋亡、分化方面發揮作用,共同影響關節軟骨的病理改變。軟骨代謝的晝夜節律已被認識很長時間,已經確定的是,骨骺生長板中軟骨細胞增殖最快的階段是清晨,這會導致生長板擴張并增加細胞外基質合成,并在中午達到峰值[9]。此外,通過體外觀察人軟骨細胞來源的細胞系,發現基因表達的自主節律,證實了生物鐘蛋白在膝關節軟骨中的表達,揭示了軟骨晝夜節律的內在本質[10-11]。軟骨細胞中基因的節律性控制可以將軟骨修復和重塑優化到一天中的最佳時間。當生物鐘基因表達出現紊亂時,軟骨的磨損與修復之間失衡,從而容易出現骨關節炎樣改變[12]。目前 Bmal1 已被發現與骨關節炎的發生、發展有關,在 Bmal1 敲除動物中發現軟骨細胞時鐘的變化可以影響軟骨的完整性。Bmal1 在骨關節炎軟骨細胞中的表達一直較低,軟骨特異性 Bmal1 敲除小鼠生長板功能受損,軟骨細胞數量減少,體長和長骨長度均下降,細胞外基質丟失明顯[5, 13-14]。這在我們的研究中也得到證實,在慢性晝夜節律紊亂的大鼠膝關節軟骨中,生物鐘基因 Bmal1 出現下調,蛋白聚糖丟失顯著,軟骨細胞凋亡增加,同時 Bmal1 的表達變化誘導了細胞周期 Wee1 基因的表達變化。
在細胞周期的分子網絡中,WEE1 作為一種 G2 期檢查點調節因子,通過介導 CDK1 的磷酸化,保護細胞核免受 CDK1-CCNB1 復合物的影響,充當有絲分裂(G2 期-M 期)的負調節因子。有研究觀察到,WEE1 可抑制含有 DNA 損傷的細胞進入有絲分裂,過表達 Wee1 可降低人卵巢癌 SKOV3 細胞凋亡,下調促凋亡蛋白 BAX 的表達,上調抗凋亡蛋白 BCL2 的表達[15-16]。為了響應 DNA 損傷,WEE1 通過 CDK1 磷酸化調節 G2/M 檢查點來負調節細胞進入有絲分裂,為細胞提供了生存優勢,允許有時間修復受損的 DNA[17-18]。WEE1 抑制消除了 G2/M 檢查點,迫使攜帶 DNA 損傷的細胞進入計劃外的有絲分裂以經歷細胞凋亡,發生有絲分裂災難[19]。CDK1 激活和失活的時間調控不僅對遺傳信息準確傳遞給子細胞至關重要,而且對確保細胞存活也至關重要。持續的 CDK1 激活對微管干擾劑誘導的細胞凋亡具有促凋亡作用[20]。CDK1-BAX 復合體構成了一個主要的死亡信號,在有絲分裂細胞周期停滯期間積累,并將主軸裝配檢查點激活與細胞死亡的開始聯系起來,并可能構成傳遞給線粒體的主要細胞質信號,從而啟動細胞死亡[21]。而 CDK1 的活性與 CCNB1 含量有密切關系,二者以復合物的形式共同調控細胞 G2 期到 M 期轉變,控制周期進展,是增殖、分裂、分化和凋亡的重要調節因子。CCNB1/CDK 復合物在 2-甲氧基雌二醇誘導的有絲分裂突變的癌細胞中上調,可通過 BCL-XL 磷酸化導致細胞凋亡[22]。此外,目前的研究證實,Bmal1 的表達下調,也可誘導組織中 Il1、Il6、Tnfa 和 Mmp13 的表達升高,血清中含量增加[23-24]。Ccnb1 和 Cdc20 的上調與外周血中 Tnfa、Il17 基因表達水平呈正相關,而巨噬細胞中 Bax/Bak 的激活會觸發 Il1 的成熟和釋放[25-26]。Il1、Il6 和 Tnfa 是參與炎癥反應的主要成分,可引起機體組織和細胞的損傷,Mmp13 是破壞軟骨基質的主要因子。說明慢性晝夜節律紊亂可通過影響生物鐘基因、細胞周期相關基因和炎癥相關基因引起軟骨蛋白聚糖的丟失,軟骨基質損傷和炎癥性改變。這些都構成了細胞衰老加快、凋亡增加和軟骨損傷的因素,在我們的研究中得到了證實。
生物鐘和細胞周期是 2 個不同又密切聯系的生物回路,有多層協調關系。二者的相關性研究多集中在腫瘤領域,關節軟骨中的實驗性研究較少。本研究發現,在晝夜節律紊亂的大鼠膝關節軟骨中,生物鐘和細胞周期共同影響了關節軟骨的病理改變,在 KOA 的發生、進展中發揮作用,這也為 KOA 分子機制研究提供了新的思路,以期開發新的治療方法,幫助患者預防或減緩 KOA 的進展。
利益沖突:所有作者聲明不存在利益沖突。
膝骨關節炎(knee osteoarthritis, KOA)是一種臨床上最常見的骨關節疾病。流行病學研究發現,長期輪班工作與髖部、腕部骨折、KOA 的風險之間存在相關性,這表明慢性晝夜節律被破壞可能會增加關節軟骨退化的易感性[1-2]。晝夜節律是一種適應性機制,由生物鐘主導調控。基本螺旋體型蛋白 1(basic helix-loop-helix ARNT like 1, BMAL1)作為生物鐘網絡的核心,是多種生理功能的重要調節劑,包括新陳代謝、睡眠、體溫、血壓和軟骨發育等。研究發現,慢性晝夜節律紊亂可引起膝關節軟骨中 Bmal1 表達下調,下游多條信號通路發生變化,是 KOA 的一個危險因素[3-4]。在軟骨特異性 Bmal1 敲除小鼠生長板中軟骨細胞增殖和存活率明顯降低,凋亡軟骨細胞的數量較高[5]。而細胞凋亡和異常分化主要受細胞周期調控,細胞周期本身也具有節律性,與生物鐘之間存在耦合關系。生物鐘基因可直接或間接控制肝臟、泌尿生殖等多個器官組織中細胞周期基因和蛋白的表達,如 WEE1 細胞周期 G2 檢測點激酶(WEE1 G2 checkpoint kinase, Wee1)、細胞周期蛋白 E 和細胞周期蛋白 B1(cyclin B1, CCNB1)等[6-8]。然而,膝關節軟骨中生物鐘和細胞周期相互影響的機制尚不完全清楚。因此,了解晝夜節律振蕩和細胞周期之間的關系,對于 KOA 的機制研究具有重要意義。
1 材料與方法
1.1 材料
1.1.1 實驗動物
40 只 5~6 周齡無特定病原體的雄性 SD 大鼠,體重 160~210 g,購自新疆醫科大學動物中心。本研究所有動物的實驗設計均獲得新疆醫科大學第一附屬醫院動物倫理委員會批準,倫理審批號:A2310-22。
1.1.2 主要試劑與儀器
番紅固綠染色試劑盒(北京索萊寶科技有限公司);聚合酶鏈反應(polymerase chain reaction, PCR)試劑盒(北京全式金生物技術有限公司);一抗[BMAL1、WEE1、細胞周期依賴性激酶 1(cyclin-dependent kinase 1, CDK1)、CCNB1、細胞凋亡調節劑 2(apoptosis regulator 2, BCL2)、BCL2 相關 X 蛋白(BCL2-associated X protein, BAX),中國武漢三鷹生物技術有限公司];二抗(山羊抗兔)、蛋白裂解液、蛋白濃度檢測試劑盒、蘇木素液和細胞凋亡檢測試劑盒(武漢塞維爾生物科技有限公司);SEDI PCR 儀(英國威泰克公司);PowerPac HC 蛋白電泳儀(美國 BIO-RAD 公司)。
1.2 方法
1.2.1 實驗分組和晝夜節律紊亂模型建立
40 只大鼠隨機分為晝夜節律正常組(正常組)、晝夜節律紊亂組(紊亂組)、Bmal1 過表達慢病毒感染的晝夜節律紊亂組(Bmal1 上調組)和 Bmal1 過表達慢病毒陰性感染的晝夜節律紊亂組(Bmal1 陰性感染組),每組 10 只。實驗期間,4 組大鼠明暗周期均為 12 h,起始 8 點開燈,20 點關燈。正常組始終保持 8 點開燈和 20 點關燈,紊亂組、Bmal1 上調組和 Bmal1 陰性感染組每隔 3 d 提前 6 h 開燈,此為 1 個循環,共進行 5 個循環,持續約 60 d,大鼠在不斷的明暗周期中發生慢性晝夜節律紊亂,見圖1。
圖1
晝夜節律紊亂模型
1.2.2 Bmal1 過表達和陰性慢病毒關節腔注射模型建立
首先制備 Bmal1 過表達和陰性慢病毒的稀釋液,滴度均為 1×108 TU/mL,置于–80°C 冰箱保存備用。晝夜節律紊亂模型開始建立的第 1 天,將 Bmal1 上調組和 Bmal1 陰性感染組的大鼠麻醉后,在左后膝關節腔內分別注射 Bmal1 過表達慢病毒和 Bmal1 過表達陰性慢病毒各 50 μL,30 d 后重復注射 1 次。
1.2.3 番紅固綠染色
模型建立后大鼠深度麻醉后放血致死,無菌條件下手術摘取膝關節,置于組織固定液中,常規脫水、透明、包埋、切片后,進行番紅固綠染色,顯微鏡下觀察軟骨病理變化。
1.2.4 原位末端轉移酶標記法(TdT-mediated dUTP nick-end labeling, TUNEL)染色
石蠟切片脫蠟至水,蛋白酶工作液覆蓋組織、37℃溫箱孵育,切片置于磷酸鹽緩沖液(phosphate buffered saline, PBS)洗滌、滴加破膜液覆蓋組織、常溫孵育,切片置于 PBS 中洗滌、滴加 buffer 覆蓋組織、常溫孵育。混合好的工作液加到組織上,于濕盒內 37℃恒溫箱孵育、PBS 洗滌、滴加 4’, 6-二脒基-2-苯基吲哚染液,避光室溫孵、PBS 中洗滌。切片甩干、抗熒光淬滅封片劑封片。熒光顯微鏡下觀察并采集圖像。
1.2.5 膝關節軟骨組織的免疫組織化學檢測
軟骨組織經多聚甲醛固定后石蠟包埋切片,脫蠟、脫水、洗片后,過氧化氫液去除內源性過氧化物酶,檸檬酸鈉和乙二胺四乙酸緩沖液進行抗原修復,一抗(BMAL1、WEE1、CDK1、CCNB1、BAX 和 BCL2)孵育過夜。第 2 天,二抗常溫孵育,二氨基聯苯胺顯色、稀鹽酸分色、蘇木素反藍,最后行脫水、透明、封片、采圖。所有染色切片通過 Image J 軟件測量陽性細胞面積進行差異性分析。
1.2.6 膝關節軟骨組織的 mRNA 表達檢測和相關性分析
主要應用 PCR 技術,將軟骨組織在加入液氮的研缽內研磨成粉,應用軟骨 RNA 柱式提取試劑盒,進行 Bmal1、Wee1、Bcl2、Cdk1、Ccnb1、Bax、白細胞介素 1(interleukin 1, Il1)、白細胞介素 6(interleukin 6, Il6)、腫瘤壞死因子(tumor necrosis factor, Tnf)和基質金屬蛋白酶 13(matrix metallopeptidase 13, Mmp13)的 RNA 提取,逆轉錄為互補 DNA 后行基因擴增。PCR 采用兩步法,擴增條件:94℃預變性 30 s,94℃變性 5 s,60℃退火和延伸 30 s,共 40 個循環。根據檢測結果的循環閾值,計算 mRNA 的相對表達量,進行差異性分析。根據實驗組中 mRNA 的相對表達量,使用 OriginPro 2021 軟件的 Correlation Plot 插件進行相關性分析。
1.2.7 膝關節軟骨組織的蛋白質印跡法檢測
將研磨后軟骨加入放射免疫沉淀測定法裂解液和蛋白磷酸酶抑制劑,在冰上提取細胞總蛋白,BCA 試劑盒測定蛋白濃度。每個蛋白樣品等量在 10% 聚丙烯酰胺凝膠上樣電泳分離,進行轉膜,5% 脫脂奶粉封閉后添加一抗(BMAL1、WEE1、CDK1、CCNB1、BAX 和 BCL2),在 4℃下過夜。PBS 溶液加上 Tween-20(PBS with Tween-20, PBST)洗滌 3 次后孵育二抗,在室溫下避光處理 1 h。PBST 洗膜 3 次,5 min/次,并用 PBS 沖洗以去除成像前的 Tween-20,化學發光成像。用 Image J 軟件進行灰度值分析。
1.3 統計學方法
應用 SPSS 23.0 軟件進行實驗數據的處理分析,統計數據服從正態分布,計量資料以均數±標準差表示,多組間比較應用單因素方差分析,多重比較采用 Tukey 法。應用 GraphPad Prism 9.0 軟件作圖,OriginPro 2021 軟件進行 Pearson 相關性分析。雙側檢驗水準 α=0.05。
2 結果
2.1 番紅固綠染色
正常組軟骨基質厚度正常,呈均勻的紅色,軟骨下骨為綠色。紊亂組軟骨基質破壞,蛋白聚糖丟失明顯,呈不均勻紅色;Bmal1 上調組軟骨基質厚度基本正常,蛋白聚糖無明顯丟失,紅色略欠均勻,軟骨下骨為綠色;Bmal1 陰性感染組軟骨出現基質破壞和蛋白聚糖丟失,呈不均質紅色。見圖2。
圖2
各組大鼠關節軟骨番紅固綠染色病理圖片(×100)
黑箭指向為蛋白聚糖
2.2 TUNEL 染色結果
凋亡的陽性細胞被染為綠色,形態、大小各異,散在不均勻地分布于正常細胞之間。與正常組和 Bmal1 上調組相比,紊亂組和 Bmal1 陰性感染組凋亡陽性細胞比率明顯升高,差異有統計學意義(P<0.001);正常組和 Bmal1 上調組相比,以及紊亂組和 Bmal1 陰性感染組相比,凋亡陽性細胞比率差異均無統計學意義(P>0.05)。見圖3 和表1。
圖3
各組大鼠關節軟骨 TUNEL 染色結果(×200)
藍色為正常軟骨細胞,綠色為凋亡的軟骨細胞。TUNEL:原位末端轉移酶標記法;merge:合并;DAPI:4’, 6-二脒基-2-苯基吲哚;spGreen:新型無毒核酸染料
2.3 各組軟骨組織中免疫組織化學分析結果
與正常組和 Bmal1 上調組相比,紊亂組和 Bmal1 陰性感染組關節軟骨中 BMAL1、WEE1 和 BCL2 蛋白的相對表達水平下降,CDK1、CCNB1 和 BAX 的蛋白相對表達水平增加,差異有統計學意義(P<0.05);正常組和 Bmal1 上調組相比,以及紊亂組和 Bmal1 陰性感染組相比,上述蛋白的相對表達水平差異均無統計學意義(P>0.05)。見表1、圖4。
圖4
細胞周期相關蛋白免疫組織化學結果(×200)
黑箭指向為染色陽性軟骨細胞
2.4 各組軟骨組織的逆轉錄 PCR 檢測和相關性分析
與正常組和 Bmal1 上調組相比,紊亂組和 Bmal1 陰性感染組關節軟骨中 Bmal1、Wee1 和 Bcl2 的 mRNA 相對表達量下降,Cdk1、Ccnb1、Bax、Il1、Il6、Tnf 和 Mmp13 的 mRNA 相對表達量增加,差異有統計學意義(P<0.05);正常組和 Bmal1 上調組相比,以及紊亂組和 Bmal1 陰性感染組相比,上述基因的 mRNA 相對表達量差異均無統計學意義(P>0.05),見表2。相關性分析顯示,Bmal1 與 Wee1、Bcl2 呈正相關,相關性系數分別為 0.84 和 0.44(P<0.01);與 Cdk1、Ccnb1、Bax 呈負相關,相關性系數分別為?0.55、?0.72、?0.41(P<0.01),見圖5。
圖5
細胞周期相關基因相關性分析
2.5 各組軟骨組織蛋白質印跡分析結果
與正常組和 Bmal1 上調組相比,紊亂組和 Bmal1 陰性感染組中 BMAL1、WEE1 和 BCL2 蛋白的相對表達水平下降,CDK1、CCNB1 和 BAX 的蛋白相對表達水平增加,差異有統計學意義(P<0.05);正常組和 Bmal1 上調組相比,以及紊亂組和 Bmal1 陰性感染組相比,上述蛋白的相對表達水平差異均無統計學意義(P>0.05),見表3 和圖6。
圖6
蛋白質印跡檢測
3 討論
本研究發現,當慢性晝夜節律紊亂引起 Bmal1 下調,給予 Bmal1 上調慢病毒干預后,關節軟骨中 Bmal1、Wee1 和 Bcl2 的表達上調,Cdk1、Ccnb1、Bax、Il1、Il6、Tnfa 和 Mmp13 的表達下調,蛋白聚糖丟失改善,軟骨細胞凋亡比率下降。我們也據此推測,Bmal1、Wee1 和 Bcl2 的升高可能會阻滯骨關節炎的進程,Cdk1、Ccnb1、Bax 的升高會加劇骨關節炎的發展。慢性晝夜節律紊亂后,生物鐘基因 Bmall 可能通過影響細胞周期相關基因,在軟骨細胞的生長、增殖、分裂和凋亡、分化方面發揮作用,共同影響關節軟骨的病理改變。軟骨代謝的晝夜節律已被認識很長時間,已經確定的是,骨骺生長板中軟骨細胞增殖最快的階段是清晨,這會導致生長板擴張并增加細胞外基質合成,并在中午達到峰值[9]。此外,通過體外觀察人軟骨細胞來源的細胞系,發現基因表達的自主節律,證實了生物鐘蛋白在膝關節軟骨中的表達,揭示了軟骨晝夜節律的內在本質[10-11]。軟骨細胞中基因的節律性控制可以將軟骨修復和重塑優化到一天中的最佳時間。當生物鐘基因表達出現紊亂時,軟骨的磨損與修復之間失衡,從而容易出現骨關節炎樣改變[12]。目前 Bmal1 已被發現與骨關節炎的發生、發展有關,在 Bmal1 敲除動物中發現軟骨細胞時鐘的變化可以影響軟骨的完整性。Bmal1 在骨關節炎軟骨細胞中的表達一直較低,軟骨特異性 Bmal1 敲除小鼠生長板功能受損,軟骨細胞數量減少,體長和長骨長度均下降,細胞外基質丟失明顯[5, 13-14]。這在我們的研究中也得到證實,在慢性晝夜節律紊亂的大鼠膝關節軟骨中,生物鐘基因 Bmal1 出現下調,蛋白聚糖丟失顯著,軟骨細胞凋亡增加,同時 Bmal1 的表達變化誘導了細胞周期 Wee1 基因的表達變化。
在細胞周期的分子網絡中,WEE1 作為一種 G2 期檢查點調節因子,通過介導 CDK1 的磷酸化,保護細胞核免受 CDK1-CCNB1 復合物的影響,充當有絲分裂(G2 期-M 期)的負調節因子。有研究觀察到,WEE1 可抑制含有 DNA 損傷的細胞進入有絲分裂,過表達 Wee1 可降低人卵巢癌 SKOV3 細胞凋亡,下調促凋亡蛋白 BAX 的表達,上調抗凋亡蛋白 BCL2 的表達[15-16]。為了響應 DNA 損傷,WEE1 通過 CDK1 磷酸化調節 G2/M 檢查點來負調節細胞進入有絲分裂,為細胞提供了生存優勢,允許有時間修復受損的 DNA[17-18]。WEE1 抑制消除了 G2/M 檢查點,迫使攜帶 DNA 損傷的細胞進入計劃外的有絲分裂以經歷細胞凋亡,發生有絲分裂災難[19]。CDK1 激活和失活的時間調控不僅對遺傳信息準確傳遞給子細胞至關重要,而且對確保細胞存活也至關重要。持續的 CDK1 激活對微管干擾劑誘導的細胞凋亡具有促凋亡作用[20]。CDK1-BAX 復合體構成了一個主要的死亡信號,在有絲分裂細胞周期停滯期間積累,并將主軸裝配檢查點激活與細胞死亡的開始聯系起來,并可能構成傳遞給線粒體的主要細胞質信號,從而啟動細胞死亡[21]。而 CDK1 的活性與 CCNB1 含量有密切關系,二者以復合物的形式共同調控細胞 G2 期到 M 期轉變,控制周期進展,是增殖、分裂、分化和凋亡的重要調節因子。CCNB1/CDK 復合物在 2-甲氧基雌二醇誘導的有絲分裂突變的癌細胞中上調,可通過 BCL-XL 磷酸化導致細胞凋亡[22]。此外,目前的研究證實,Bmal1 的表達下調,也可誘導組織中 Il1、Il6、Tnfa 和 Mmp13 的表達升高,血清中含量增加[23-24]。Ccnb1 和 Cdc20 的上調與外周血中 Tnfa、Il17 基因表達水平呈正相關,而巨噬細胞中 Bax/Bak 的激活會觸發 Il1 的成熟和釋放[25-26]。Il1、Il6 和 Tnfa 是參與炎癥反應的主要成分,可引起機體組織和細胞的損傷,Mmp13 是破壞軟骨基質的主要因子。說明慢性晝夜節律紊亂可通過影響生物鐘基因、細胞周期相關基因和炎癥相關基因引起軟骨蛋白聚糖的丟失,軟骨基質損傷和炎癥性改變。這些都構成了細胞衰老加快、凋亡增加和軟骨損傷的因素,在我們的研究中得到了證實。
生物鐘和細胞周期是 2 個不同又密切聯系的生物回路,有多層協調關系。二者的相關性研究多集中在腫瘤領域,關節軟骨中的實驗性研究較少。本研究發現,在晝夜節律紊亂的大鼠膝關節軟骨中,生物鐘和細胞周期共同影響了關節軟骨的病理改變,在 KOA 的發生、進展中發揮作用,這也為 KOA 分子機制研究提供了新的思路,以期開發新的治療方法,幫助患者預防或減緩 KOA 的進展。
利益沖突:所有作者聲明不存在利益沖突。