直接使用速度信息和角度信息驅動網格細胞吸引子對環境編碼的方法,抗干擾能力較差且不具有仿生性。針對這一問題,本文提出一種基于感知速度與感知角度的網格野計算模型。其特點在于,通過對視覺流進行解碼處理獲得視覺速度,對速度細胞建模并解碼獲得本體速度,對視覺速度和本體速度進行融合求得感知速度信息;利用加入興奮性連接的一維環狀模型模擬頭朝向細胞的放電機制,使機器人以仿生的方式獲取當前的感知角度信息。最后,融合速度和角度兩種感知信息實現對網格細胞吸引子模型的驅動。對所提模型進行實驗驗證,結果表明該模型可以實現網格細胞周期性六邊形放電野模式以及精確的路徑積分功能。研究成果為仿海馬認知機制的機器人認知地圖構建方法研究奠定了基礎。
引用本文: 于乃功, 馮慧, 廖詣深, 鄭相國. 一種基于感知速度與感知角度的網格野計算模型. 生物醫學工程學雜志, 2020, 37(5): 863-874. doi: 10.7507/1001-5515.201911058 復制
版權信息: ?四川大學華西醫院華西期刊社《生物醫學工程學雜志》版權所有,未經授權不得轉載、改編
引言
生理學研究表明,海馬結構是大鼠進行環境認知的關鍵腦區[1]。海馬結構中存在多種空間細胞,如頭朝向細胞、速度細胞、網格細胞和位置細胞等。1984 年 Ranck 等在大鼠的前下托發現了對頭部方向具有強烈感知作用的神經元細胞,該類神經元細胞的放電率會隨著大鼠頭部朝向的改變而變化,因此被命名為頭朝向細胞[2-3]。1998 年,O’Keefe 等對速度細胞的存在提出了猜想,并認為速度細胞的放電率和大鼠的運動速度呈正相關關系。2015 年 Kropff 等[4]通過實驗證實了速度細胞的存在,并引起了學術界對空間認知過程中速度信號的編碼規則與作用機制的思考。研究表明,通過整合速度細胞提供的信息,大鼠可以實現對運動速度的感知,這為大鼠構建精確的認知地圖提供了生理學依據。2005 年 Hafting 等[5]通過變換實驗箱大小和形狀的實驗,發現了對空間位置具有強烈放電作用的網格細胞,網格細胞會在大鼠所探索的空間中發生穩定的周期性放電,并形成六邊形的網格野覆蓋大鼠遍歷的整個空間環境[6-7]。網格野可以起到度量大鼠所遍歷空間的距離和角度的作用,從而指導大鼠確定空間坐標,實現自我定位,完成對空間環境的認知。振蕩干涉模型[8-11]和吸引子模型[12-13]是兩種經典的網格細胞計算模型。
行為學研究表明,動物能夠利用自運動線索保持相對環境重要位置的矢量關系,并將空間中的各個位點的信息進行計算和整合,使這種矢量關系不斷得到更新,這一過程被稱為路徑積分[14-15]。研究表明,網格細胞在大鼠大腦中充當路徑整合的角色[16]。速度和角度是路徑積分過程中的主要輸入信息,并且該過程是網格細胞網格野形成的主要機制。其中,頭朝向細胞提供感知方向信息[17],速度細胞提供不含方向信息的運動速度信號。然而,在大鼠腦內直接參與環境認知過程的速度信息并非僅由速度細胞提供,而是由多種信號共同編碼得到的一種感知速度。直接參與路徑積分過程的感知速度是由包括視覺因素在內的多種因素影響而產生的。波士頓大學 Raudies 等[18]的改變視覺-空間輸入的實驗以及清華大學謝康寧[19]的控制大鼠運動的實驗都證明了上述結論。
先前的網格細胞網格野計算模型,直接使用由編碼器解算的速度值驅動網格細胞吸引子,沒有對速度細胞進行建模研究,這種方法缺乏生理學依據,并且單純依靠編碼器信息驅動網格細胞吸引子的方法抗干擾能力相對較差[13]。2018 年本團隊[14]建立了一種頭朝向細胞計算模型,該計算模型給定了頭朝向細胞間的調整內核,但未對細胞之間的權重連接給出分析,模型不夠完善且對頭朝向信號的跟隨不夠精確。針對以上問題,本文根據海馬結構中的空間細胞的生理學研究[20]以及感知速度的獲取模式研究,首先,對速度細胞進行建模并且提出了一種融合視覺速度和本體速度從而獲得感知速度的計算方法;其次,利用加入興奮性連接的一維環狀細胞模型模擬了頭朝向細胞的放電機制,提出了一種感知角度的獲取方法;最后,利用感知速度和感知角度驅動網格細胞吸引子模型,實現網格細胞對空間環境的編碼。
1 模型架構
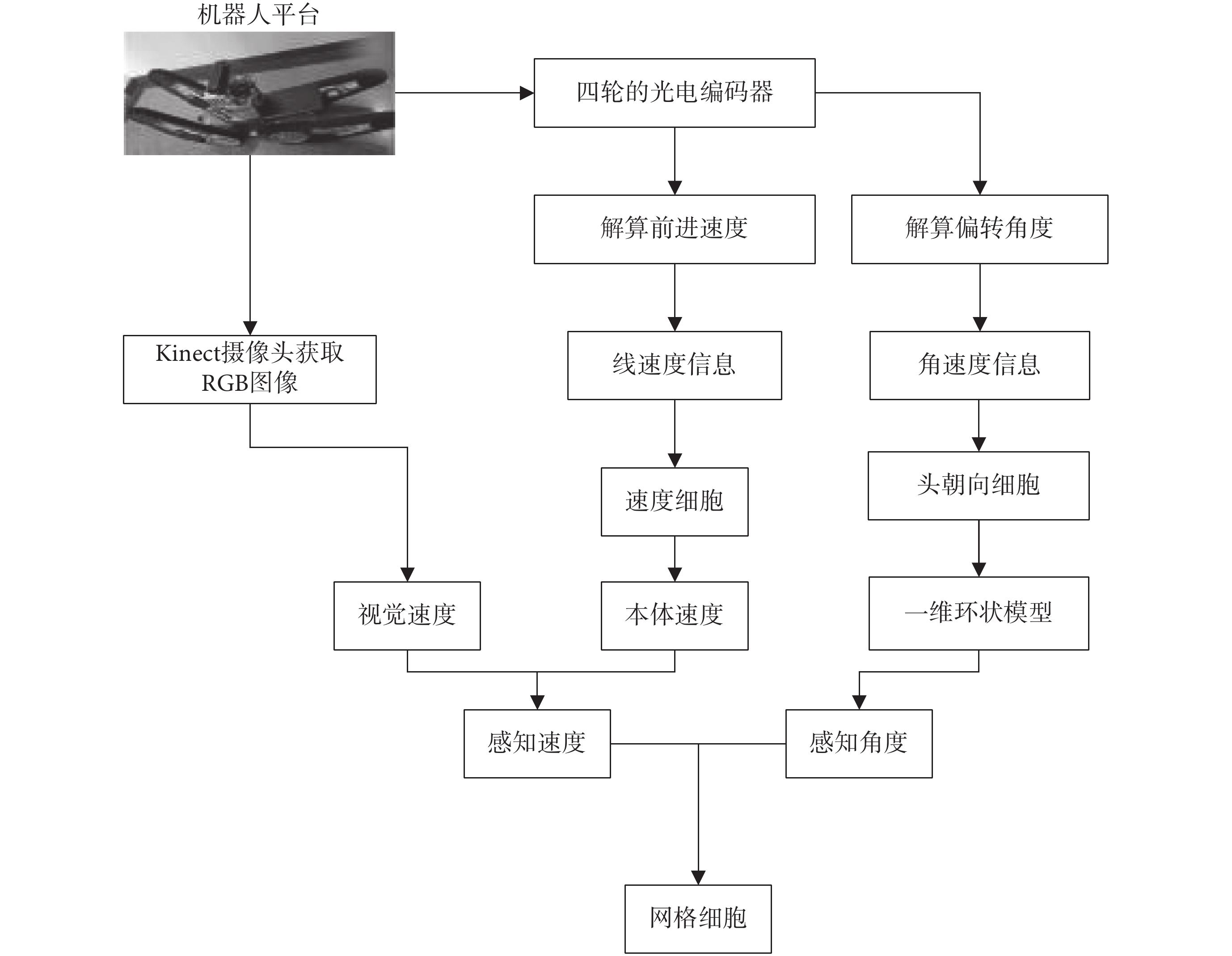
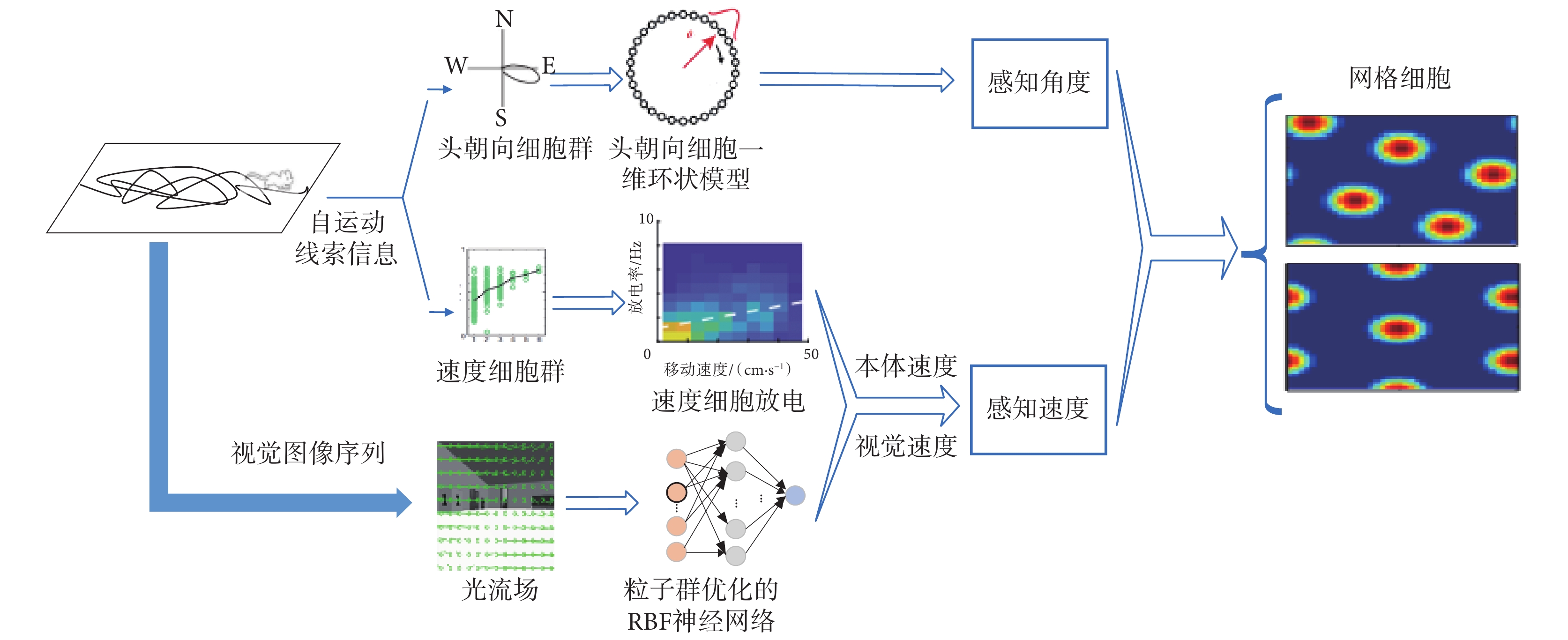
本文所構建的基于感知速度與感知角度的網格野計算模型如圖 1 所示。首先,攝像頭采集環境的圖像信息,編碼器采集機器人的速度信息與方向信息。然后,融合視覺速度和本體速度以仿生的方式獲取感知速度信息。接著,將頭朝向細胞建模成加入興奮性連接的一維環狀模型,以仿生的方式獲取感知角度信息。最后,將感知速度信息和感知角度信息輸入網格細胞二維連續吸引子模型,驅動網格細胞板上興奮活動包的移動,實現網格細胞對空間環境的編碼以及路徑積分功能。
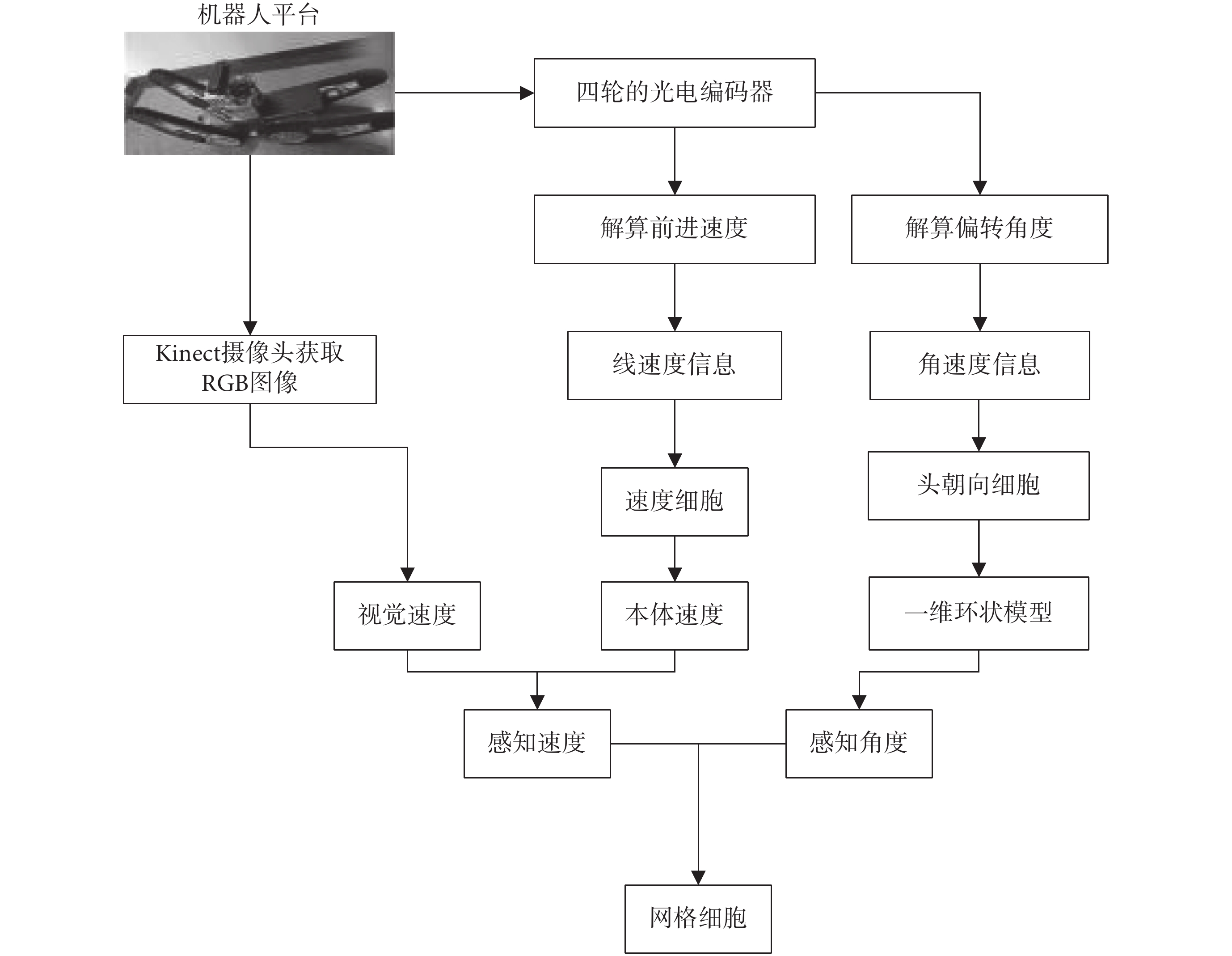 圖1
基于感知速度與感知角度的網格野計算模型
Figure1.
Grid field calculation model based on perceived speed and perceived angle
圖1
基于感知速度與感知角度的網格野計算模型
Figure1.
Grid field calculation model based on perceived speed and perceived angle
2 感知速度的獲取
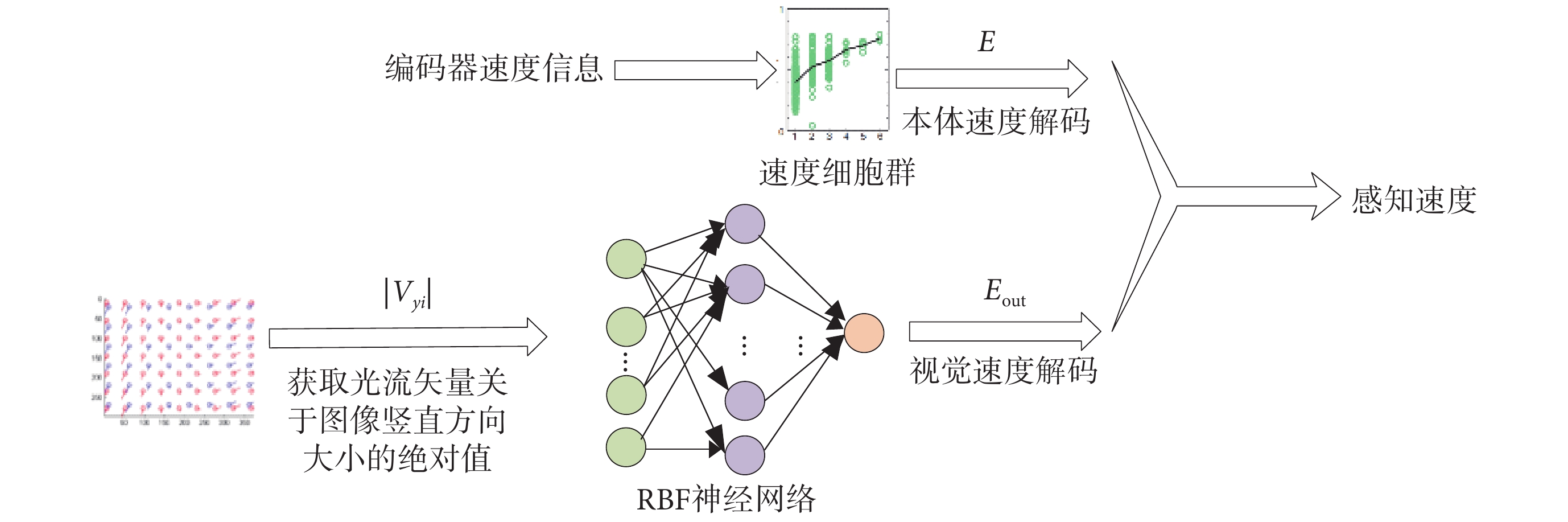
感知速度是由多種信號協同編碼產生的,且感知速度與實際速度之間不可避免地會存在誤差。影響感知速度的主要因素有:視覺流和本體感覺。視網膜圖像的連續變化稱為視覺流,這種連續變化會引起生物體對自身速度的估計。本體感覺指的是由速度細胞構成的速度碼系統,該系統獲取不含方向信息的運動速率作為本體速度[21]。基于此,本文提出一種基于速度細胞和視覺信息的感知速度求解方法。圖 2 為感知速度獲取原理示意圖。
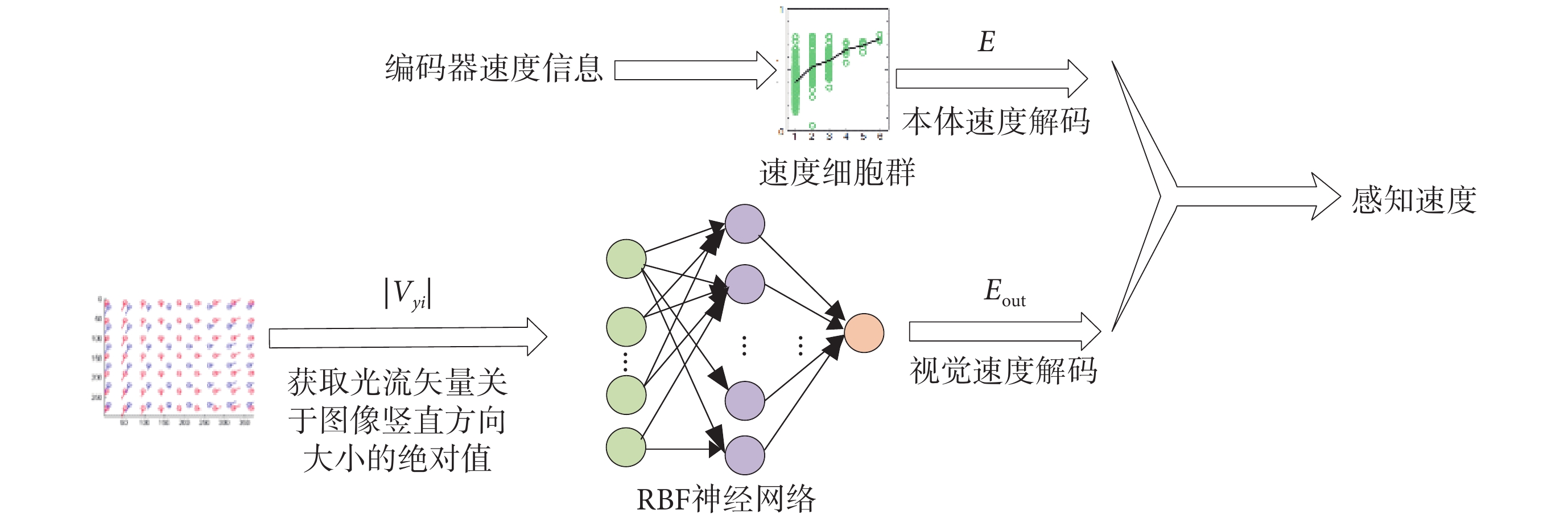 圖2
感知速度獲取原理示意圖
Figure2.
Schematic diagram of the principle of perceived speed acquisition
圖2
感知速度獲取原理示意圖
Figure2.
Schematic diagram of the principle of perceived speed acquisition
2.1 基于光流法的視覺速度獲取
將光流算法應用到本模型中,采用經典的 Farneback 光流算法[22]計算連續圖像間的光流場,關于光流算法的具體內容本文不再進行闡述。根據亮度恒定假設、圖像梯度恒定假設以及空間一致性計算所有圖像序列的光流矢量,光流矢量關于圖像豎直方向分量大小的絕對值記為 。
。
 |
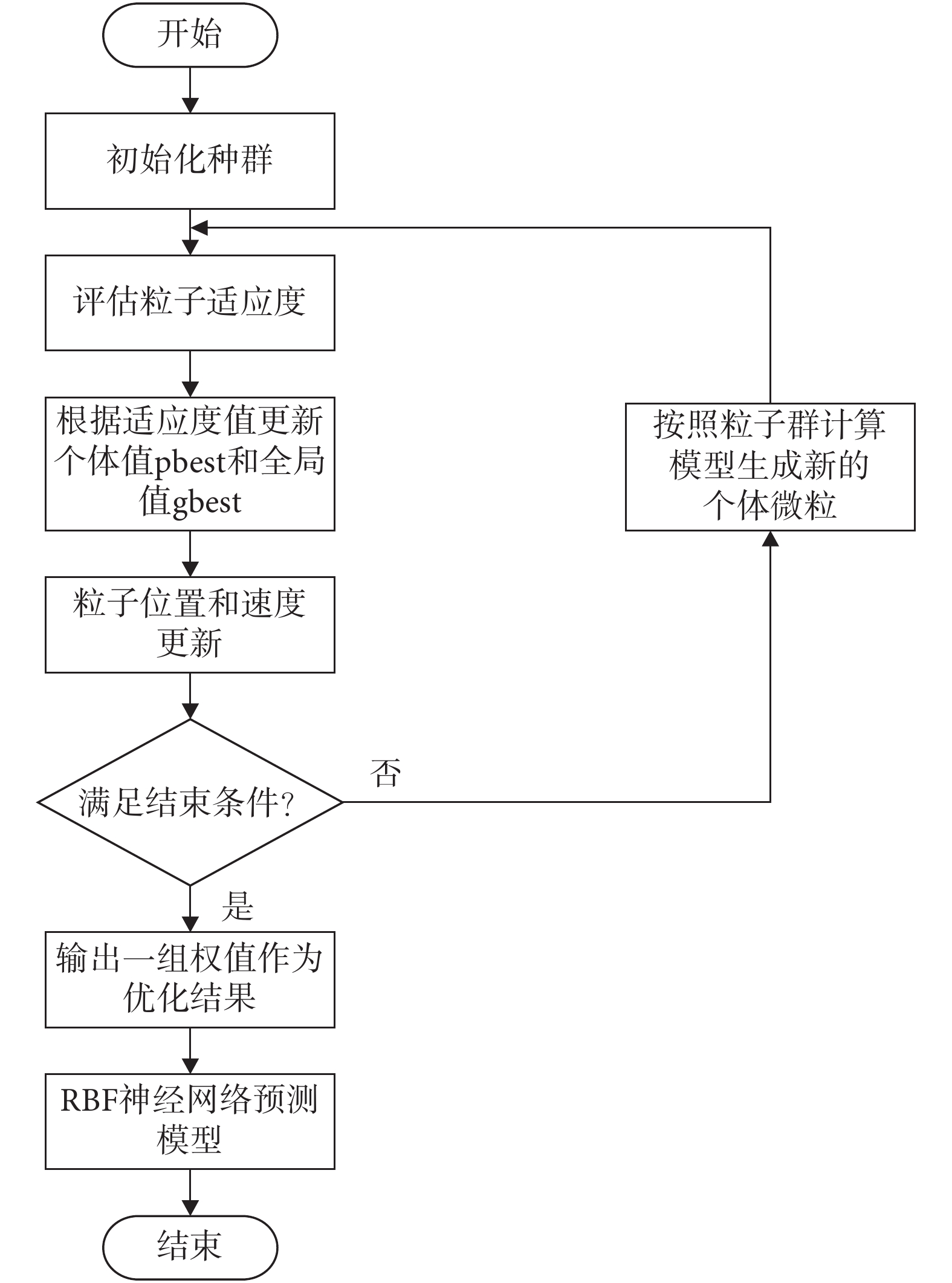
采用粒子群優化的徑向基函數(radial basis function,RBF)神經網絡[23-24]作為通過視覺估計線速度的算法。為避免陷入局部最小值,采用粒子群算法對神經網絡權值進行優化,取光流矢量關于圖像豎直方向分量大小的絕對值 作為神經網絡的輸入樣本,以每一幀圖像拍攝時機器人實際運行的線速度作為輸出樣本,對神經網絡進行訓練。權值優化算法流程圖如圖 3 所示。
作為神經網絡的輸入樣本,以每一幀圖像拍攝時機器人實際運行的線速度作為輸出樣本,對神經網絡進行訓練。權值優化算法流程圖如圖 3 所示。
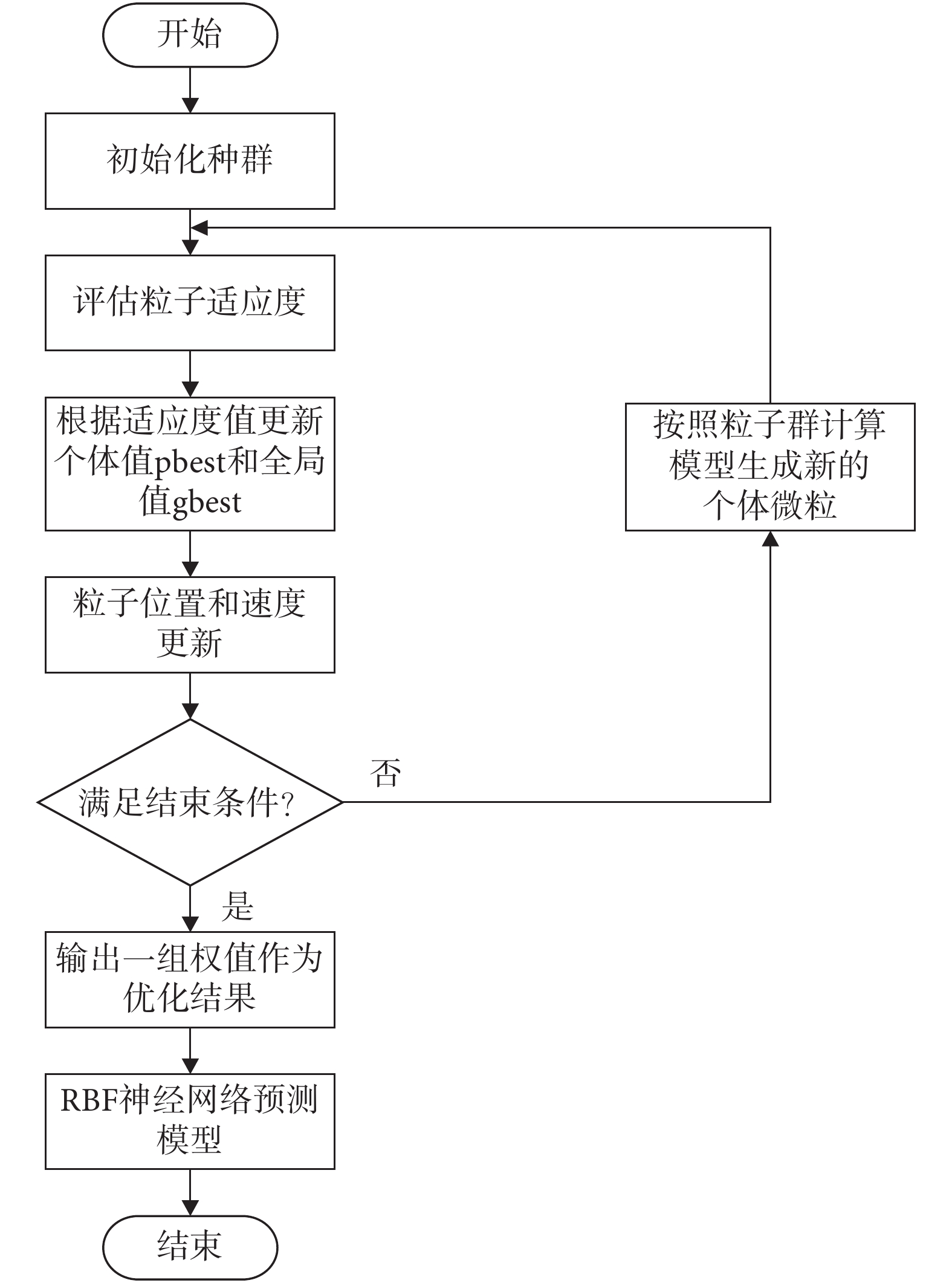 圖3
粒子群優化 RBF 神經網絡權值算法流程圖
Figure3.
Algorithm flow chart of particle swarm optimization of the weight of RBF neural network
圖3
粒子群優化 RBF 神經網絡權值算法流程圖
Figure3.
Algorithm flow chart of particle swarm optimization of the weight of RBF neural network
2.2 基于速度細胞模型的本體速度獲取
生理學研究表明,速度細胞的放電率與大鼠的運動速度呈正相關關系,并且當大鼠處于靜止狀態時速度細胞也會產生放電現象[4, 25]。基于生理學實際,本文構建了如下的速度細胞模型。
定義速度細胞的放電率公式為
 |
 |
 |
式(2)中  代表第
代表第  個速度細胞的放電率,
個速度細胞的放電率, 、
、 大于零,其中,
大于零,其中, 為速度細胞感知速度的基礎偏置量,
為速度細胞感知速度的基礎偏置量, 為速度細胞對速度值的感知比例。
為速度細胞對速度值的感知比例。 小于零,保證了速度細胞對運動速度具有良好的跟隨性。如式(3)所示,
小于零,保證了速度細胞對運動速度具有良好的跟隨性。如式(3)所示, 為增加了擾動之后速度細胞所感知到的速度信息。
為增加了擾動之后速度細胞所感知到的速度信息。 為速度細胞感知速度的擾動值,如式(4)所示擾動信號服從標準正態分布。
為速度細胞感知速度的擾動值,如式(4)所示擾動信號服從標準正態分布。
為使本文所構建的速度細胞模型更符合生理學依據,定義速度細胞置信度為大鼠的運動速度和放電率之間的 Pearson 相關系數。
 |
式(5)中  為所有速度細胞放電率的平均值,
為所有速度細胞放電率的平均值, 為運動速度的平均值,
為運動速度的平均值, 為速度細胞群中速度細胞的數量。公式計算所得
為速度細胞群中速度細胞的數量。公式計算所得  為速度細胞的置信度,閾值范圍為 0.94~0.97,當
為速度細胞的置信度,閾值范圍為 0.94~0.97,當  小于閾值時檢查速度細胞模型,并從中選擇離群值淘汰,得到精確度更高的速度細胞群,從而使得歸一化后計算的速度細胞放電率的數學期望值更具可靠性。對滿足閾值條件的速度細胞群做如下處理。
小于閾值時檢查速度細胞模型,并從中選擇離群值淘汰,得到精確度更高的速度細胞群,從而使得歸一化后計算的速度細胞放電率的數學期望值更具可靠性。對滿足閾值條件的速度細胞群做如下處理。
首先,計算單個速度細胞的放電率在速度細胞群放電率中的權值  。
。
 |
式(6)中  為剔除離群值之后速度細胞的數量,然后對所有速度細胞的放電率做歸一化處理。
為剔除離群值之后速度細胞的數量,然后對所有速度細胞的放電率做歸一化處理。
 |
式(7)中  為歸一化處理后第
為歸一化處理后第  個速度細胞的放電率,
個速度細胞的放電率, 為速度細胞群中速度細胞放電率的標準差,
為速度細胞群中速度細胞放電率的標準差, 的計算公式如式(8)所示。
的計算公式如式(8)所示。
 |
最后根據式(9)求解速度細胞群放電率的數學期望  ,即所有速度細胞放電率的加權平均值。
,即所有速度細胞放電率的加權平均值。
 |
2.3 感知速度的求解
當視覺流的連續變化受到附加速度的影響較大時,會由于視覺對比度產生對自身速度的嚴重錯估。當編碼器信息受到路面環境的輕微擾動時,會使得對本體速度的估計產生偏差。因此在求解感知速度  的時候,采用公式(10)對感知速度進行求解。
的時候,采用公式(10)對感知速度進行求解。
 |
式(10)中, 和
和  分別為速度細胞放電率的數學期望
分別為速度細胞放電率的數學期望  和神經網絡的輸出
和神經網絡的輸出  轉化至同一量綱下的結果,
轉化至同一量綱下的結果, 代表權值系數,
代表權值系數, 代表調整閾值,當
代表調整閾值,當  和
和  之間的差值大于
之間的差值大于  時,判斷視覺對比度引起了對自身速度的嚴重錯估,令感知速度為
時,判斷視覺對比度引起了對自身速度的嚴重錯估,令感知速度為  ;當
;當  和
和  之間的差值小于等于
之間的差值小于等于  時,判斷視覺速度估算正確,同時為了減小由于擾動導致的本體速度估值的偏差,令
時,判斷視覺速度估算正確,同時為了減小由于擾動導致的本體速度估值的偏差,令  和
和  通過權值系數
通過權值系數  求解得到的加權平均值作為感知速度
求解得到的加權平均值作為感知速度  。
。
3 感知角度的獲取
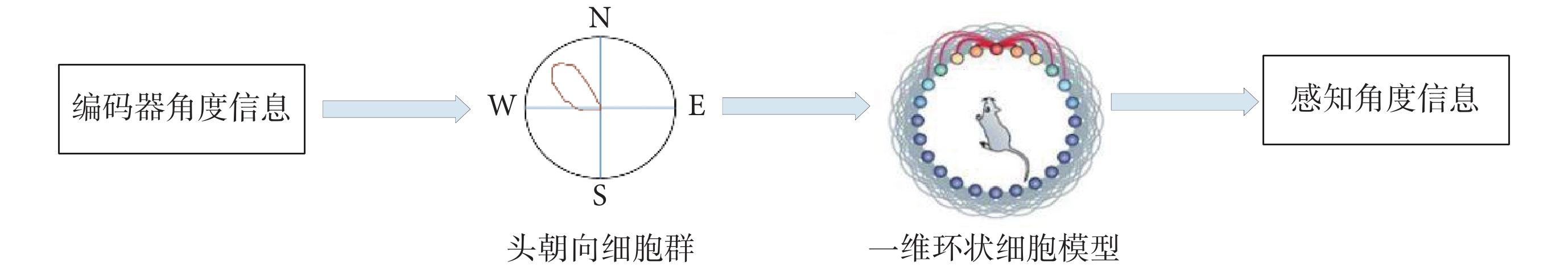
圖 4 為感知角度獲取原理示意圖,通過一維環狀細胞模型對頭朝向細胞進行建模,獲取環狀模型上興奮活動包的位置實現感知角度的獲取。
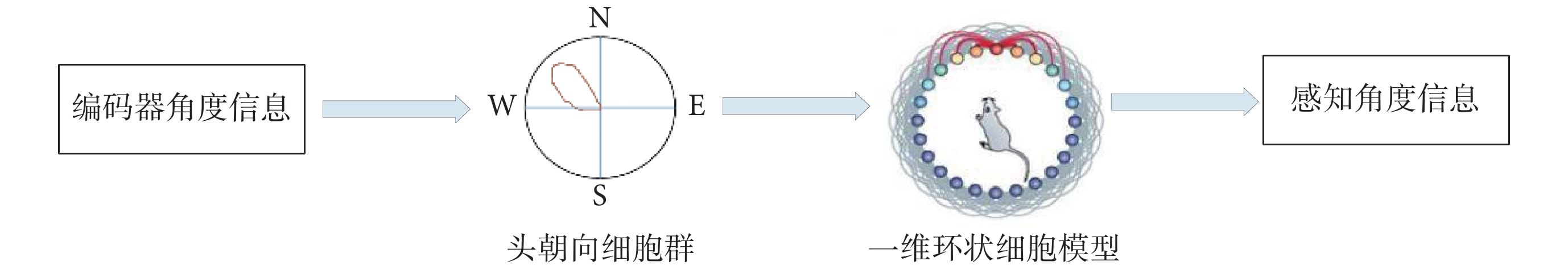 圖4
感知角度獲取原理示意圖
Figure4.
Schematic diagram of the principle of perceived angle acquisition
圖4
感知角度獲取原理示意圖
Figure4.
Schematic diagram of the principle of perceived angle acquisition
3.1 頭朝向細胞的建模
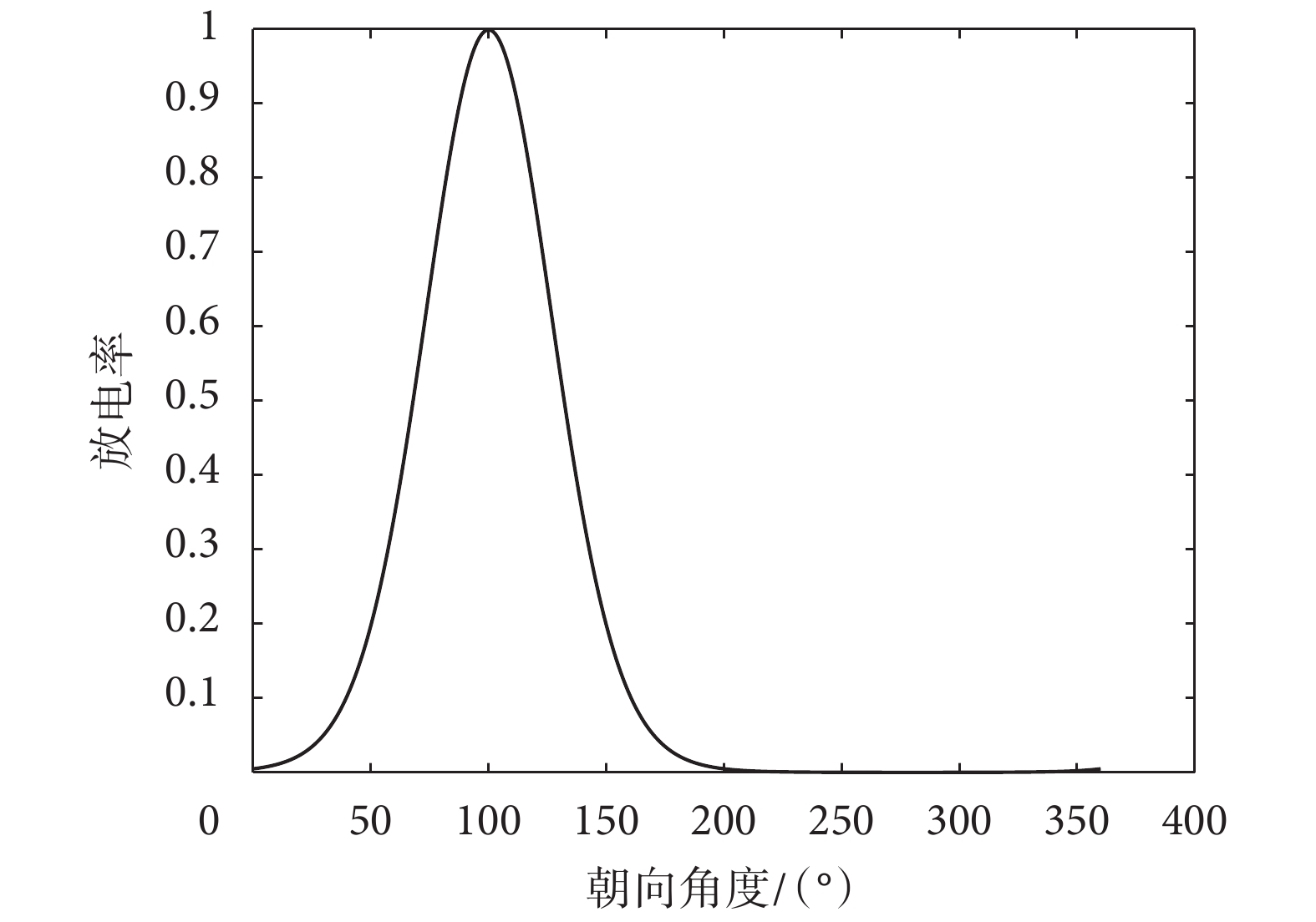
頭朝向細胞具有如下放電特性[26-27]:以水平方向為起點,細胞的放電強度隨著大鼠頭朝向角度的增加而增加,當大鼠的頭朝向到達該頭朝向細胞的最佳頭部方向時,放電率達到峰值,然后跟隨頭朝向角度的增大而逐漸偏離最佳方向,并且放電強度也降低[14]。根據頭朝向細胞的生理學研究事實,定義頭朝向細胞的放電率公式如式(11)所示。
 |
式(11)中, 代表頭朝向細胞
代表頭朝向細胞  的放電率,
的放電率, 代表當前頭朝向角度,
代表當前頭朝向角度, 代表該頭朝向細胞的最優方向,K 決定放電率曲線的銳度,
代表該頭朝向細胞的最優方向,K 決定放電率曲線的銳度, 和
和  分別決定頭朝向細胞的放電基準值和放電率峰值大小。根據公式(11),設定
分別決定頭朝向細胞的放電基準值和放電率峰值大小。根據公式(11),設定  等于 0、
等于 0、 等于 0.01 以及 K 等于 4.53 的優先方向為 100° 的頭朝向細胞的放電特性如圖 5 所示。
等于 0.01 以及 K 等于 4.53 的優先方向為 100° 的頭朝向細胞的放電特性如圖 5 所示。
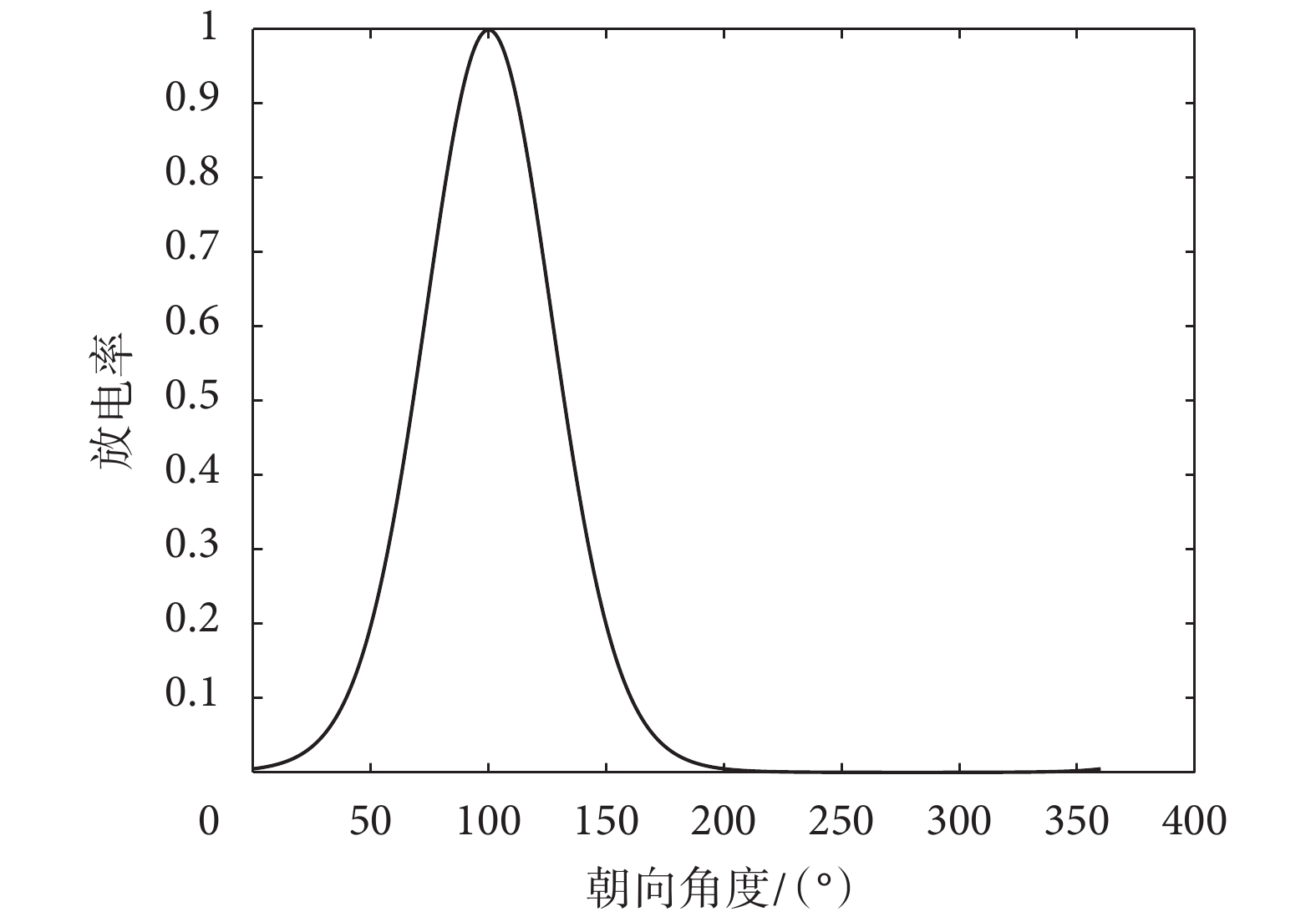 圖5
頭朝向細胞放電特性示意圖
Figure5.
Schematic diagram of discharge characteristics of head direction cell
圖5
頭朝向細胞放電特性示意圖
Figure5.
Schematic diagram of discharge characteristics of head direction cell
3.2 一維環狀頭朝向細胞的建模
將順序排列的頭朝向細胞首尾連接形成一個封閉的環形,第  個頭朝向細胞對應的最優方向為
個頭朝向細胞對應的最優方向為  ,并且可以表示為相對主朝向
,并且可以表示為相對主朝向  的角偏移量,
的角偏移量, 的計算公式如式(12)所示。
的計算公式如式(12)所示。
 |
其中  代表頭朝向細胞的編號,
代表頭朝向細胞的編號, 代表一維環狀模型中頭朝向細胞的個數。
代表一維環狀模型中頭朝向細胞的個數。
根據頭朝向細胞的放電特性,可知在環狀細胞模型中將會出現一個高斯帽狀的興奮活動包,隨著頭朝向角度的變化,興奮活動包會在環狀細胞模型上移動。獲取興奮活動包在環狀模型上的位置,即可得知當前的頭朝向信息。
隨著興奮活動包在環狀細胞模型上長時間移動,興奮活動包會逐漸擴散,將導致從環狀細胞模型上獲取的角度信息與真實角度信息產生誤差。因此,本文提出一種頭朝向細胞之間加入興奮連接的神經網絡模型,頭朝向細胞之間的連接強度計算公式如式(13)所示[26]。
 |
其中, 、
、 為常系數,
為常系數, 是興奮性連接基準值,
是興奮性連接基準值, 是興奮參數權值。
是興奮參數權值。 是常系數,
是常系數, 、
、 分別表示頭朝向細胞
分別表示頭朝向細胞  和頭朝向細胞
和頭朝向細胞  在環狀模型的位置坐標,從公式(13)可以看出頭朝向細胞之間的興奮性連接強度只和相互連接的頭朝向細胞之間的角度差有關。
在環狀模型的位置坐標,從公式(13)可以看出頭朝向細胞之間的興奮性連接強度只和相互連接的頭朝向細胞之間的角度差有關。
由興奮性連接產生的頭朝向細胞的放電變化量為
 |
式(14)中, 代表頭朝向細胞之間的興奮性連接權值,
代表頭朝向細胞之間的興奮性連接權值, 代表頭朝向細胞
代表頭朝向細胞  的放電率,
的放電率, 代表一維環狀細胞模型中頭朝向細胞的個數。興奮性傳遞之后,將所有頭朝向細胞的放電率與 0 進行比較,以保證所有頭朝向細胞的放電率都不小于零,然后做歸一化處理,其數學表達式如式(15)和(16)所示。
代表一維環狀細胞模型中頭朝向細胞的個數。興奮性傳遞之后,將所有頭朝向細胞的放電率與 0 進行比較,以保證所有頭朝向細胞的放電率都不小于零,然后做歸一化處理,其數學表達式如式(15)和(16)所示。
 |
 |
4 網格細胞網格野計算模型
在沒有任何外界輸入信息的環境中,網格細胞能夠根據自運動線索中的感知速度和感知角度進行精確的路徑積分,網格細胞的放電活動被認為能夠形成空間環境的度量地圖[28]。本文在 Burak 等[13]提出的網格細胞模型基礎上進行了改進,提出了一種基于感知速度與感知角度的網格野計算模型,本文模型中網格細胞接收的輸入信息不再是未經處理的編碼器信息,而是將編碼器信息以前面所提仿生的方式進行處理,獲得更具有生理學依據且抗干擾性更強的感知速度和感知角度。網格細胞網格野計算模型結構圖如圖 6 所示。
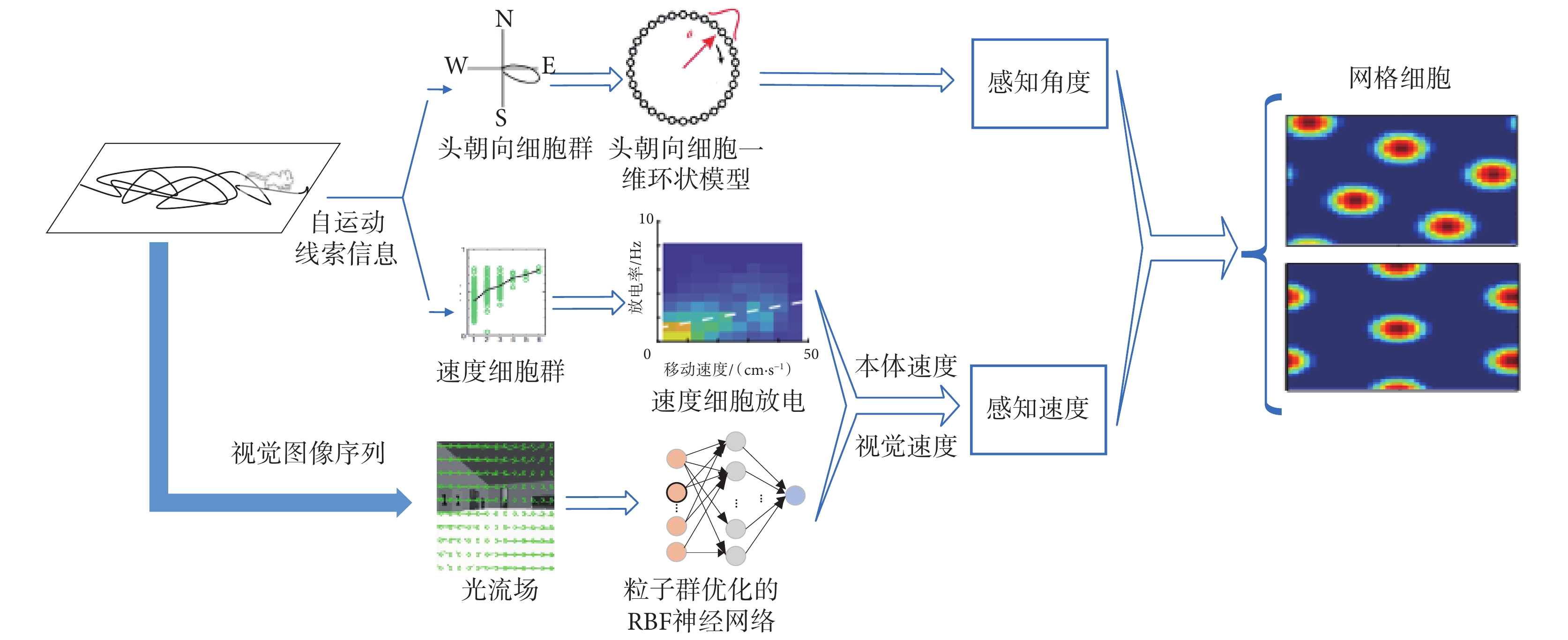 圖6
網格細胞網格野計算模型結構圖
Figure6.
Structure diagram of grid field calculation model
圖6
網格細胞網格野計算模型結構圖
Figure6.
Structure diagram of grid field calculation model
網格細胞吸引子受兩方面的驅動:一是網格細胞之間的抑制性遞歸輸入;二是來自上游皮層的興奮性投射信息。使用二維連續吸引子模型對網格細胞進行建模,每個神經元對應一個吸引子[14]。隨著大鼠在空間環境中的不斷探索,遍歷的范圍也逐漸增大,然而表示空間相對位置的神經元不能無限增加,從而會涉及到吸引子網絡的邊界問題,所以將神經面板的上下邊界和左右邊界相互連接,形成二維環狀吸引子模型[29]。二維的神經面板中網格細胞的神經元動力學等式為
 |
其中, 是神經元相應的時間常量,傳遞函數
是神經元相應的時間常量,傳遞函數  是一個非線性整流函數,當
是一個非線性整流函數,當  時
時  ,當
,當  時
時  ,
, 時刻當前位置神經元
時刻當前位置神經元  的狀態為
的狀態為  ,
, 是神經面板上神經元
是神經面板上神經元  到神經元
到神經元  的連接權值,
的連接權值, 是神經面板上周圍神經元到神經元
是神經面板上周圍神經元到神經元  的抑制性遞歸輸入,
的抑制性遞歸輸入, 是來自上游皮層的興奮性投射信息。假定神經面板的長度為
是來自上游皮層的興奮性投射信息。假定神經面板的長度為  ,那么整個神經網絡有
,那么整個神經網絡有  個神經元,每個神經元
個神經元,每個神經元  都有一個優先朝向,并且優先朝向信息由頭朝向細胞產生的感知角度確定。感知角度信息確定輸出權值的改變方向,網格細胞的遞歸連接權值為
都有一個優先朝向,并且優先朝向信息由頭朝向細胞產生的感知角度確定。感知角度信息確定輸出權值的改變方向,網格細胞的遞歸連接權值為
 |
 |
權值矩陣的分布將形成中間高兩邊低的墨西哥帽狀形狀,其中間位置為  ,
, 為對
為對  方向的感知速度進行積分之后的周期性重置距離。在所有的實驗中設定
方向的感知速度進行積分之后的周期性重置距離。在所有的實驗中設定  ,
, ,其中
,其中  為神經面板上網格的周期。
為神經面板上網格的周期。
下面給出感知速度與感知角度對網格細胞吸引子的作用機制。定義  時刻大鼠的頭朝向角度為
時刻大鼠的頭朝向角度為  ,沿著該方向上的感知速度為
,沿著該方向上的感知速度為  ,那么沿著方向
,那么沿著方向  的感知速度為
的感知速度為
 |
對速度進行積分得到  方向的位移為
方向的位移為
 |
分別定義放電周期  和相位
和相位  來決定感知速度的輸入模式,其中
來決定感知速度的輸入模式,其中  決定網絡速度響應的增益,放電周期
決定網絡速度響應的增益,放電周期  為一固定值,那么對
為一固定值,那么對  方向的感知速度進行積分之后的周期性重置距離為
方向的感知速度進行積分之后的周期性重置距離為
 |
定義網格細胞所接收的興奮性輸入為
 |
 |
 代表了對輸入信息的耦合并通過感知角度信息確定了網格細胞吸引子的移動方向。
代表了對輸入信息的耦合并通過感知角度信息確定了網格細胞吸引子的移動方向。
5 實驗驗證
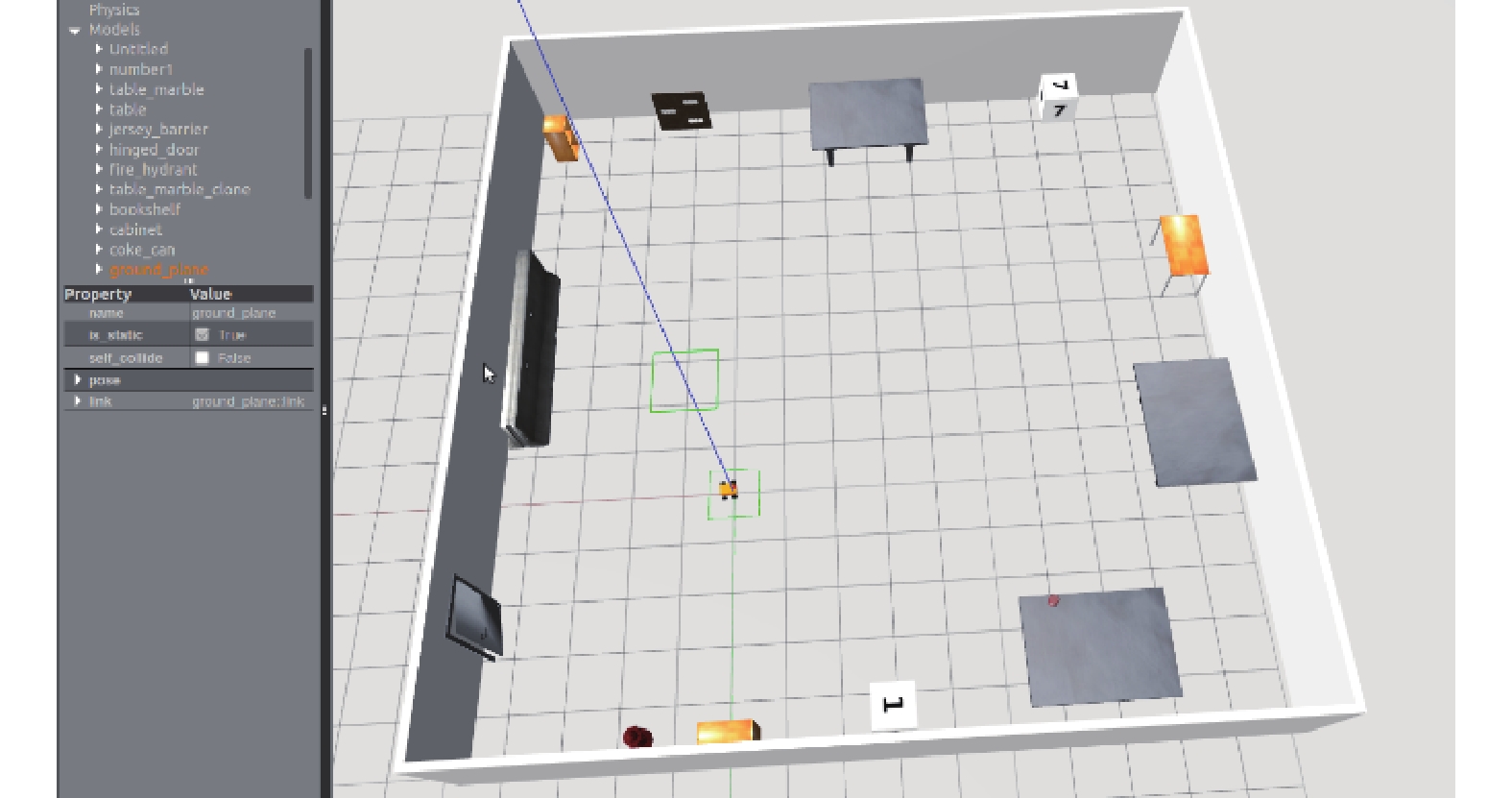
利用仿真實驗和物理實驗對本文所構建的模型進行驗證。為驗證所構建的感知角度和感知速度模型的合理性以及網格野計算模型的有效性和魯棒性,采用 ROS 自帶的 Gazeboard 仿真器搭建一個 15 m*15 m 的虛擬環境并控制機器人對該環境進行探索,采集里程計信息及環境 RGB 圖像進行實驗驗證。通過實際機器人平臺進行了走廊環境路徑積分實驗,驗證了由感知速度和感知角度驅動的網格細胞路徑積分功能的精確性。仿真實驗環境如圖 7 所示。
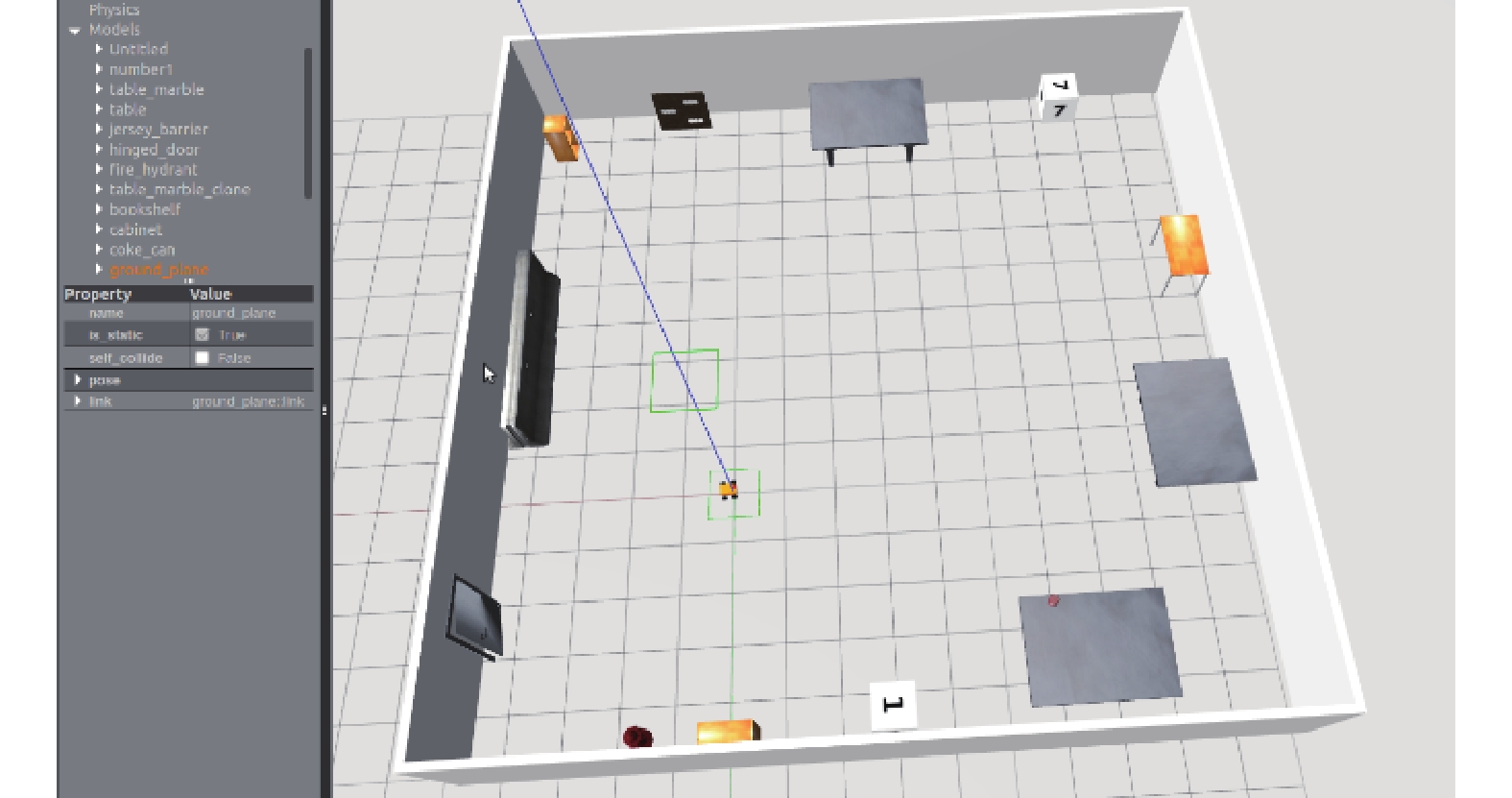 圖7
仿真實驗界面
Figure7.
Simulation experiment interface
圖7
仿真實驗界面
Figure7.
Simulation experiment interface
5.1 感知速度模型實驗驗證
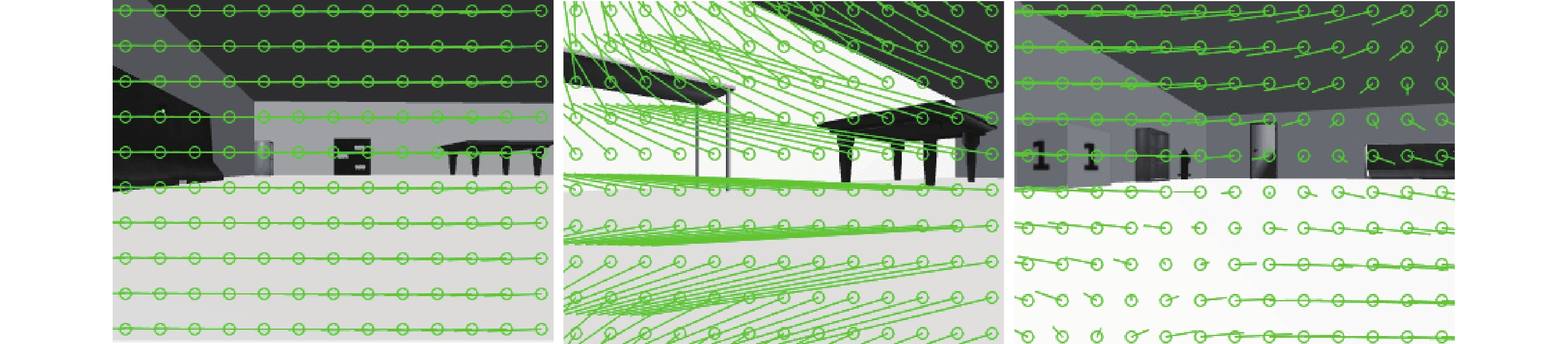
利用機器人平臺在虛擬環境下探索 30 min,實時獲取機器人移動過程中的里程計信息及 RGB 圖像,計算得到每幀圖像拍攝時機器人的移動速度及連續圖像間的光流矢量。采用 Farneback 光流算法計算得到的虛擬環境下的光流場效果如圖 8 所示。設計 RBF 神經網絡作為通過視覺估計線速度的算法,取光流矢量關于圖像豎直方向分量大小的絕對值作為輸入信息,以每一幀圖像拍攝時機器人實際運行的線速度作為輸出樣本,并采用粒子群算法對神經網絡權值以及 RBF 的中心和寬度進行優化。設置粒子群迭代次數為 250 次,種群規模為 30,粒子維度 12,加速度因子  。設定 Farneback 光流算法計算光流場的分辨率為 10*130,即 RBF 神經網絡有 130 個輸入節點,1 個輸出節點,通過正交最小二乘法計算得到隱含層中心數為 22。設定神經網絡學習率為 0.05,均方誤差為 0.000 3,迭代次數為 240 次,以上參數的設定值為多次實驗后得到的經驗值。
。設定 Farneback 光流算法計算光流場的分辨率為 10*130,即 RBF 神經網絡有 130 個輸入節點,1 個輸出節點,通過正交最小二乘法計算得到隱含層中心數為 22。設定神經網絡學習率為 0.05,均方誤差為 0.000 3,迭代次數為 240 次,以上參數的設定值為多次實驗后得到的經驗值。
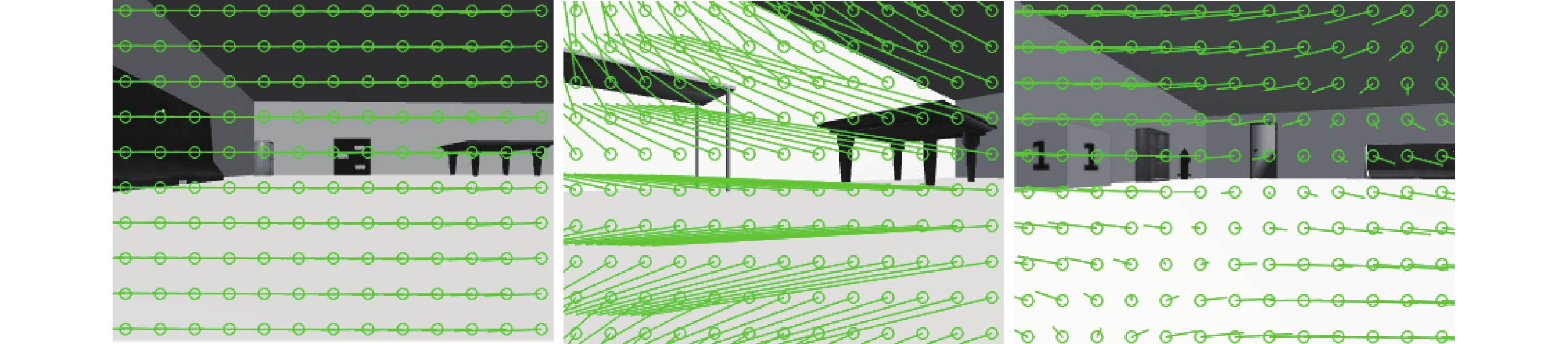 圖8
仿真環境下的圖像光流場效果圖
Figure8.
Optical flow vector of image in simulation environment
圖8
仿真環境下的圖像光流場效果圖
Figure8.
Optical flow vector of image in simulation environment
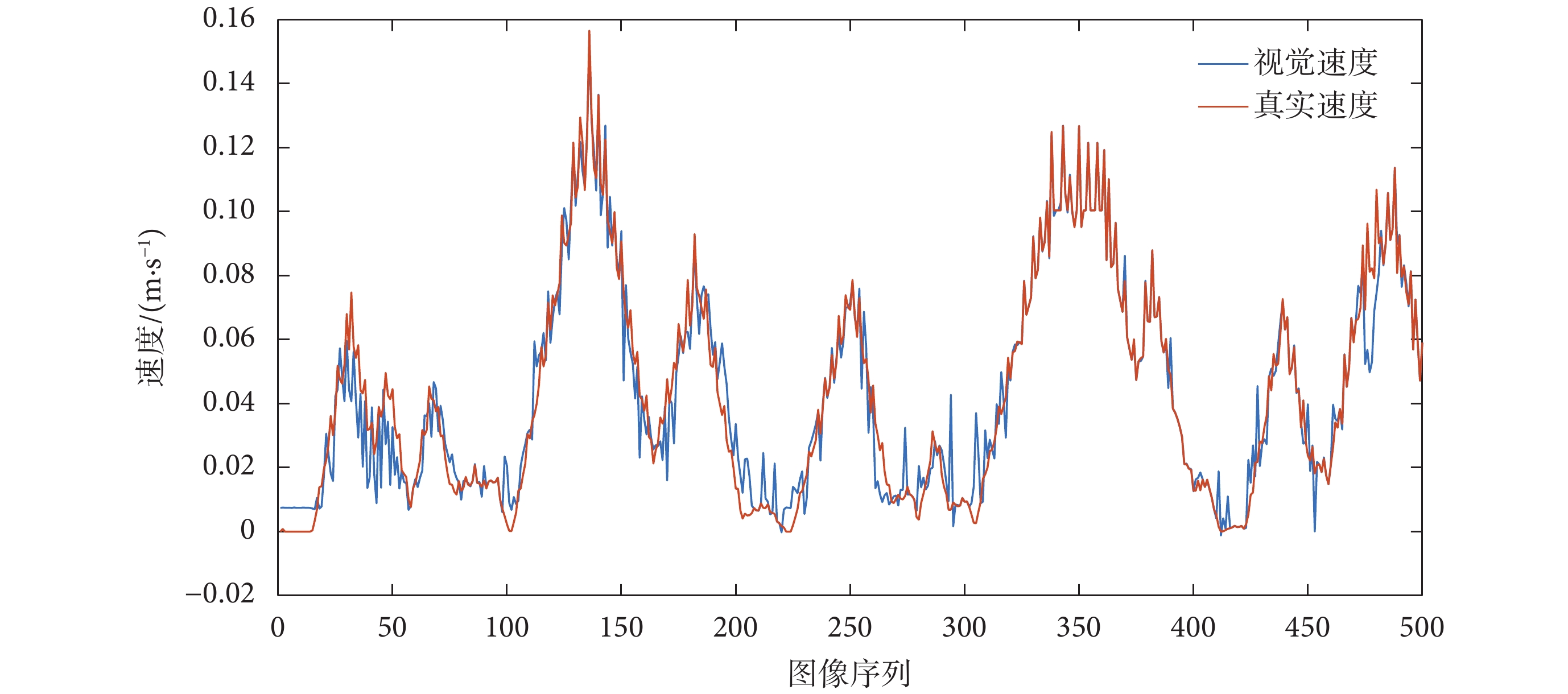
對 RBF 神經網絡訓練的過程是一個參數尋優的過程,將粒子群中的每一個體分量映射到網絡中的權值,對每一個體對應的神經網絡輸入機器人采集的連續圖像幀并進行訓練,網絡中權值的優化過程是一個反復迭代的過程,迭代的終止條件是最大迭代次數。對訓練后的網絡進行測試,由 RBF 神經網絡得出的視覺速度與機器人的實際運動速度對比圖如圖 9 所示。
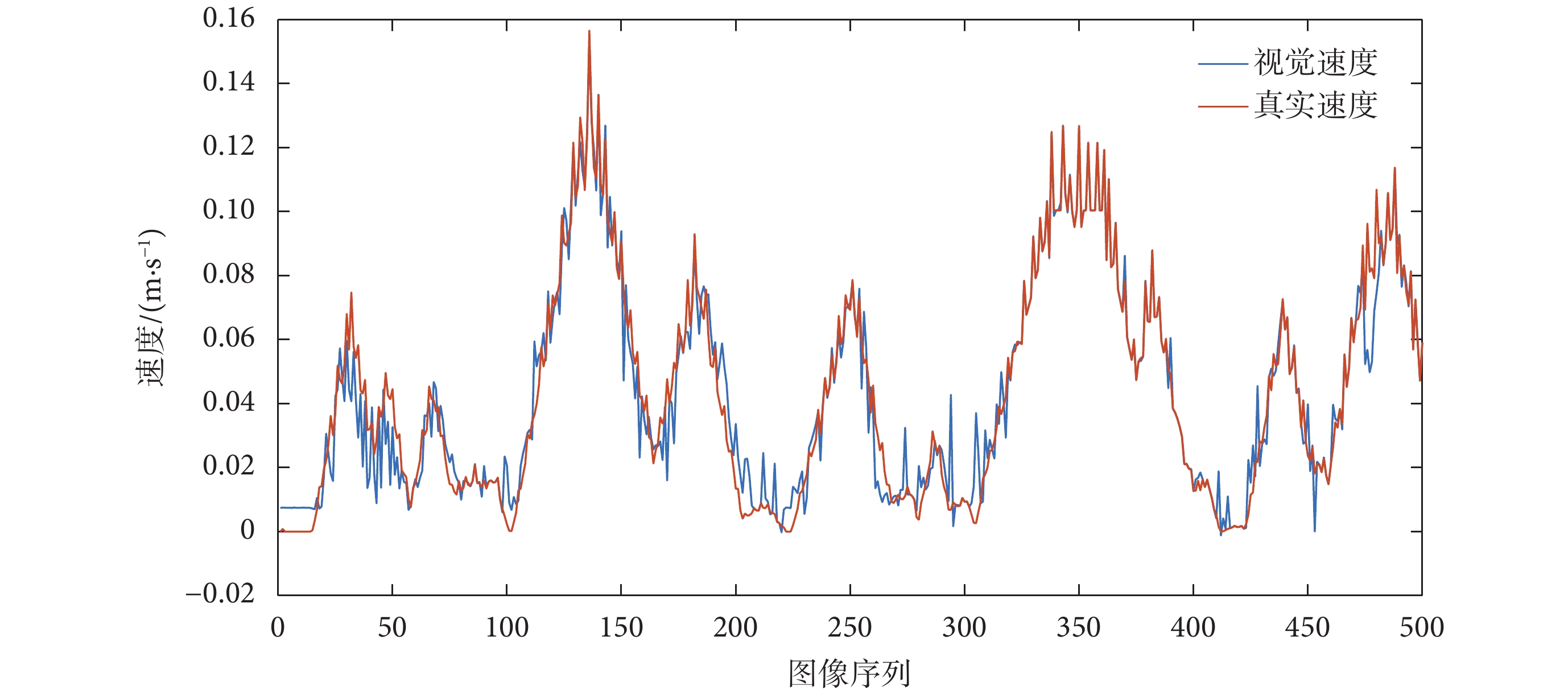 圖9
視覺速度和真實速度對比圖
Figure9.
Comparison chart of visual speed and real speed
圖9
視覺速度和真實速度對比圖
Figure9.
Comparison chart of visual speed and real speed
對 2.2 節所提的速度細胞數學模型進行實驗驗證,為保證模型的合理性與準確性,對速度細胞放電率參數設定如下: 、
、 、
、 、
、 均設定為服從正態分布,其中,
均設定為服從正態分布,其中, 服從均值為 4、方差為 2 的正態分布;
服從均值為 4、方差為 2 的正態分布; 服從均值為 1.5、方差為 3 的正態分布;
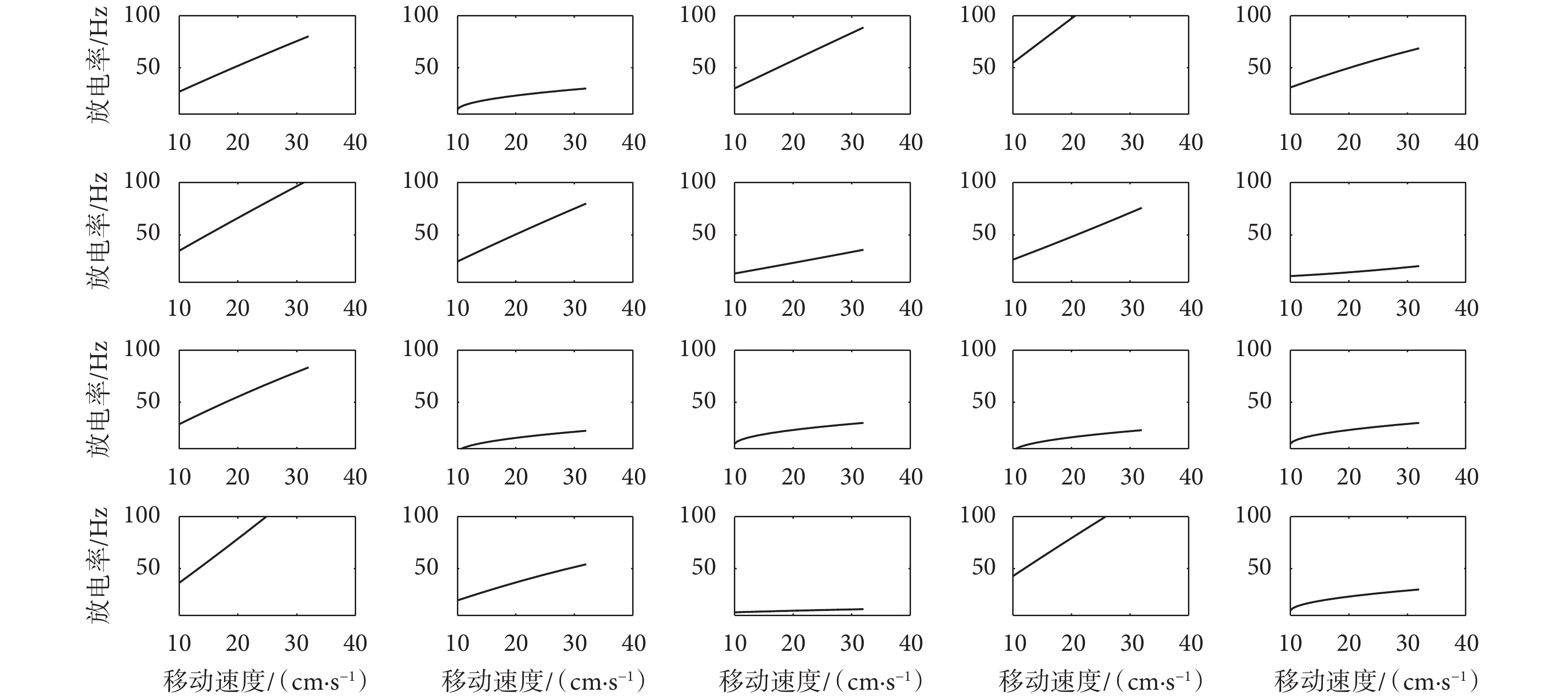
服從均值為 1.5、方差為 3 的正態分布; 服從均值為-0.005、方差為 0.001 的正態分布;Vr 服從均值為 0、方差為 1 的正態分布。同時,設定速度細胞個數為 20 個。對速度細胞數學模型進行實驗驗證,得到如圖 10 所示的不同參數值下的速度細胞放電率對比圖。圖中所示的速度細胞的放電率與運動速度之間的相關系數均大于規定的閾值,表明了本文模型所構建的速度細胞的可靠性,且符合速度細胞放電率與大鼠的運動速度呈正相關關系這一生理學現象。
服從均值為-0.005、方差為 0.001 的正態分布;Vr 服從均值為 0、方差為 1 的正態分布。同時,設定速度細胞個數為 20 個。對速度細胞數學模型進行實驗驗證,得到如圖 10 所示的不同參數值下的速度細胞放電率對比圖。圖中所示的速度細胞的放電率與運動速度之間的相關系數均大于規定的閾值,表明了本文模型所構建的速度細胞的可靠性,且符合速度細胞放電率與大鼠的運動速度呈正相關關系這一生理學現象。
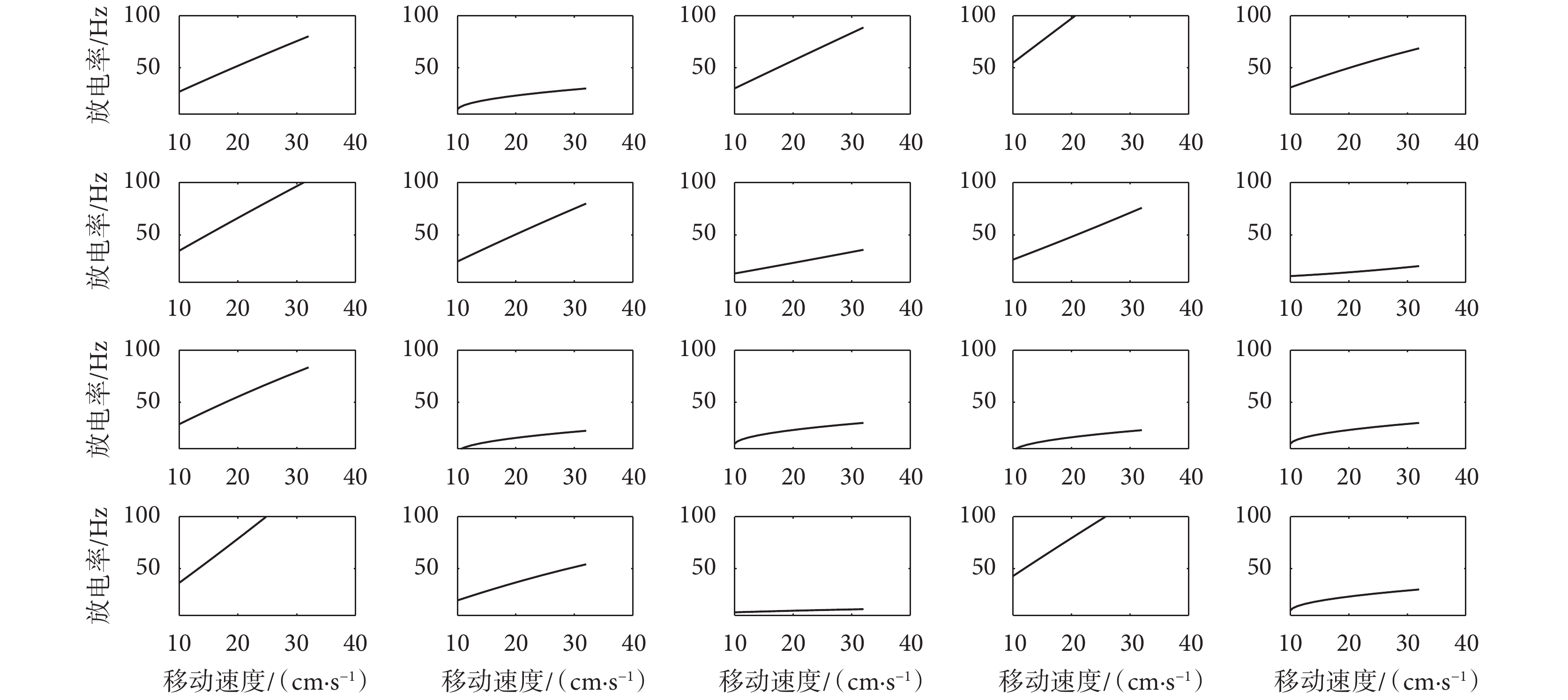 圖10
不同參數值下的速度細胞放電率圖
Figure10.
Speed cell firing rate map under different parameter values
圖10
不同參數值下的速度細胞放電率圖
Figure10.
Speed cell firing rate map under different parameter values
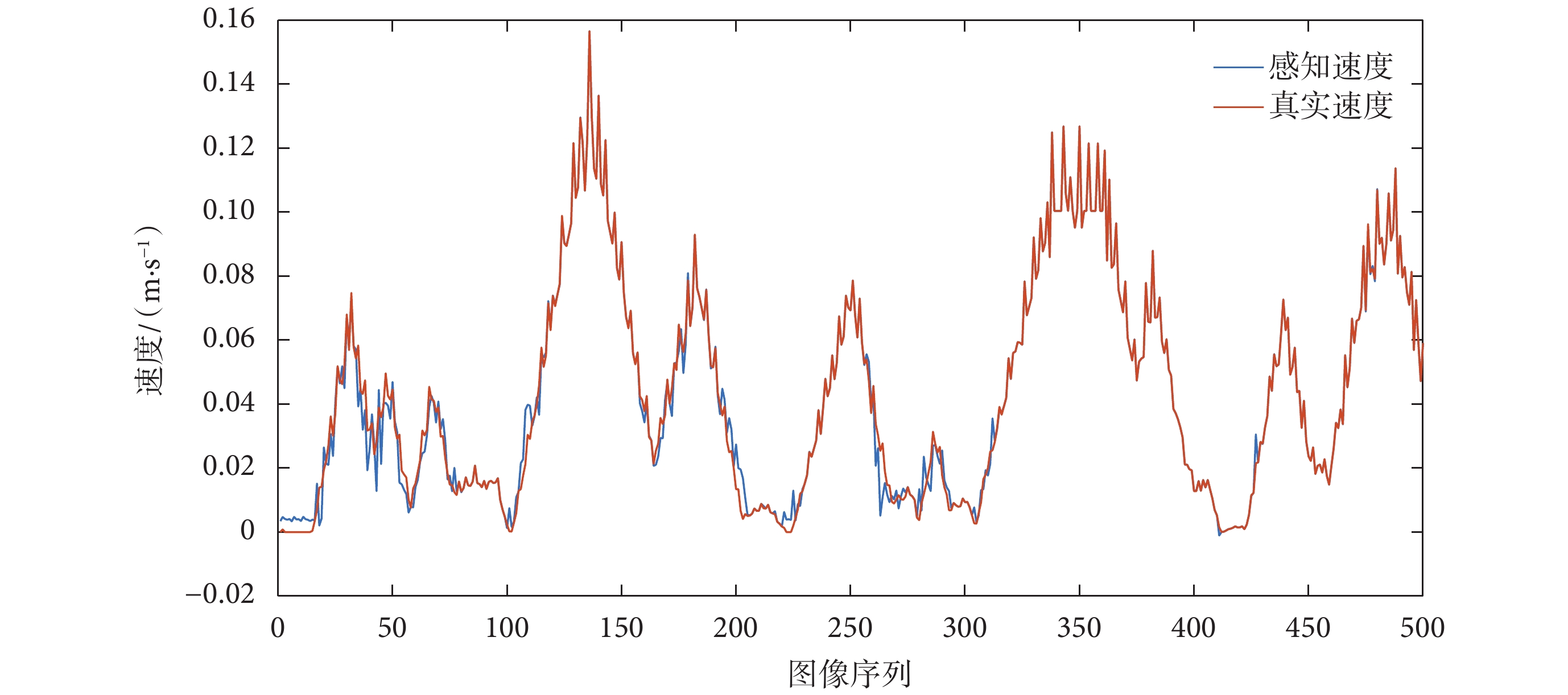
為模擬真實環境中依靠編碼器信息解算速度值存在干擾與偏差這一現象,將仿真環境中得到的真實速度加入動態的輕微擾動,模型中令擾動服從標準正態分布。根據 2.2 節所提算法,速度細胞群對加入擾動之后的速度信息進行處理并解碼獲得本體速度。根據本文融合算法,將本體速度與通過 RBF 神經網絡獲得的視覺速度進行加權平均,得到感知速度  ,感知速度與真實速度對比圖如圖 11 所示。
,感知速度與真實速度對比圖如圖 11 所示。
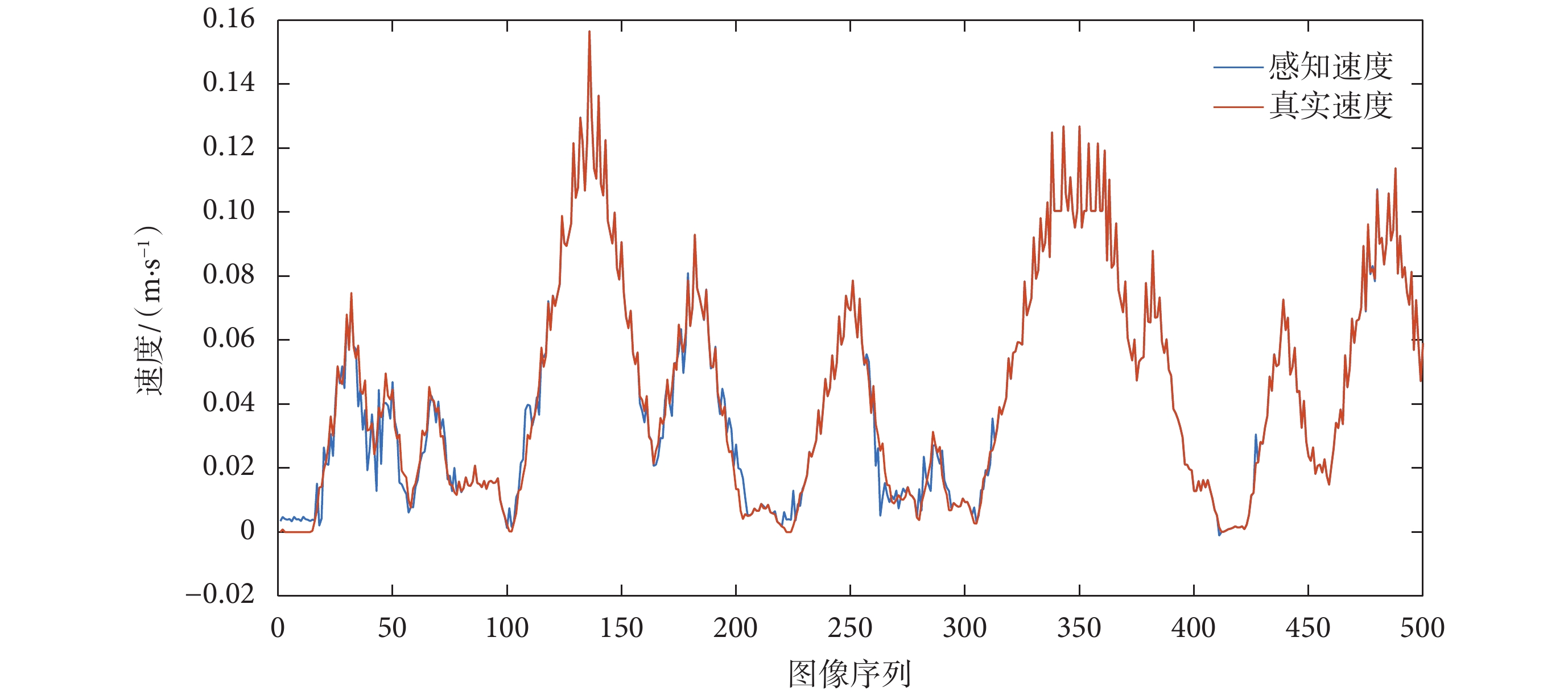 圖11
感知速度和真實速度對比圖
Figure11.
Comparison chart of perceived speed and real speed
圖11
感知速度和真實速度對比圖
Figure11.
Comparison chart of perceived speed and real speed
從圖 9 和 圖 11 可以看出,基于光流解算得到的視覺速度基本可以實現對當前運動速度的估計,基于視覺速度和本體速度融合得到的感知速度與真實速度基本接近,同時又不完全依賴于機器人本身所攜帶的編碼器對速度的測量。表明所構建的感知速度獲取模型符合直接參與環境認知過程的速度信息是多種信號協同編碼的結果這一生理學實際,且感知速度與實際速度之間不可避免地存在誤差。
5.2 感知角度模型實驗驗證
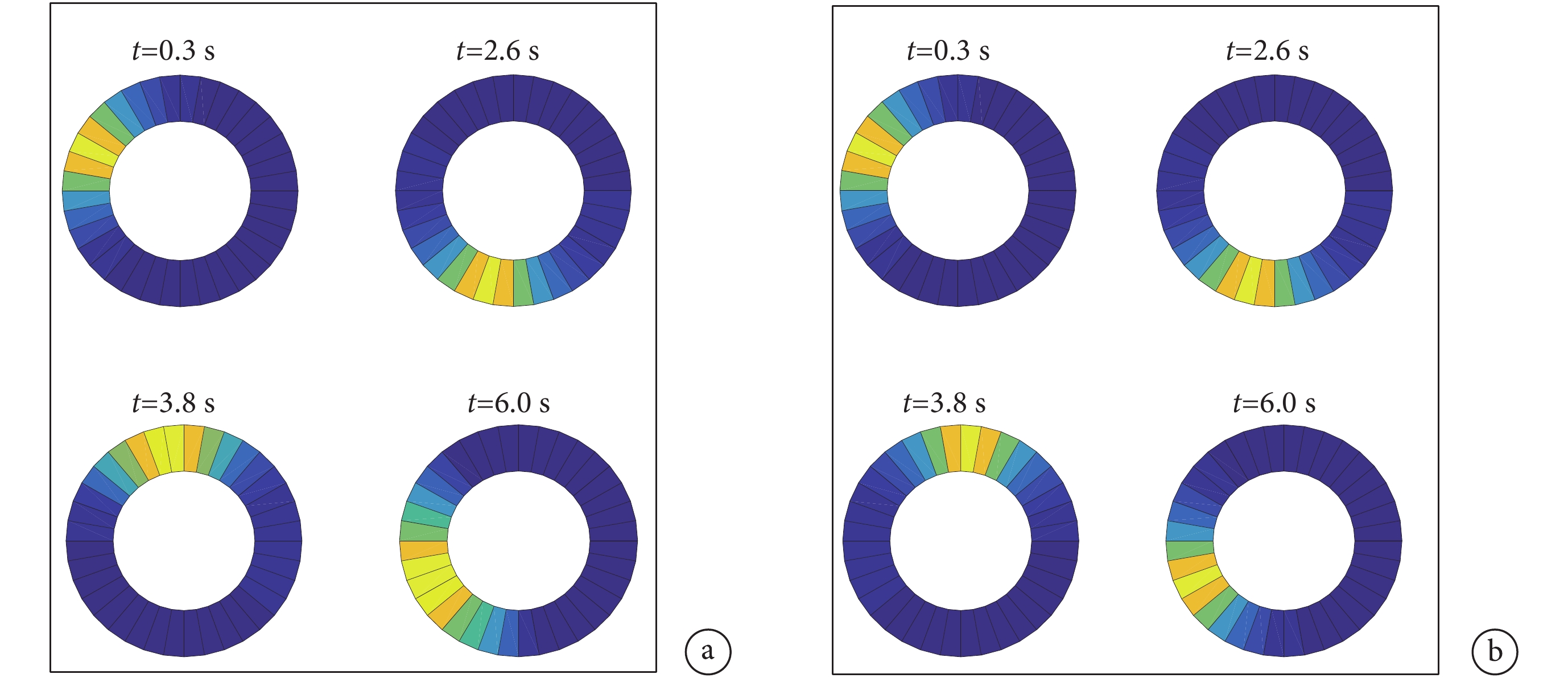
本文中將頭朝向細胞建模成一維環狀模型,根據頭朝向細胞的放電特性可知在環狀細胞模型上將會產生一個高斯帽狀的興奮活動包。隨著興奮活動包在環狀細胞模型上的長時間移動興奮活動包會逐漸擴散,這會導致從環狀細胞模型上獲取的感知角度信息與真實的角度信息之間產生誤差。對未加入興奮連接的環狀細胞模型進行仿真實驗得到的興奮活動包擴散的效果如圖 12a 所示。
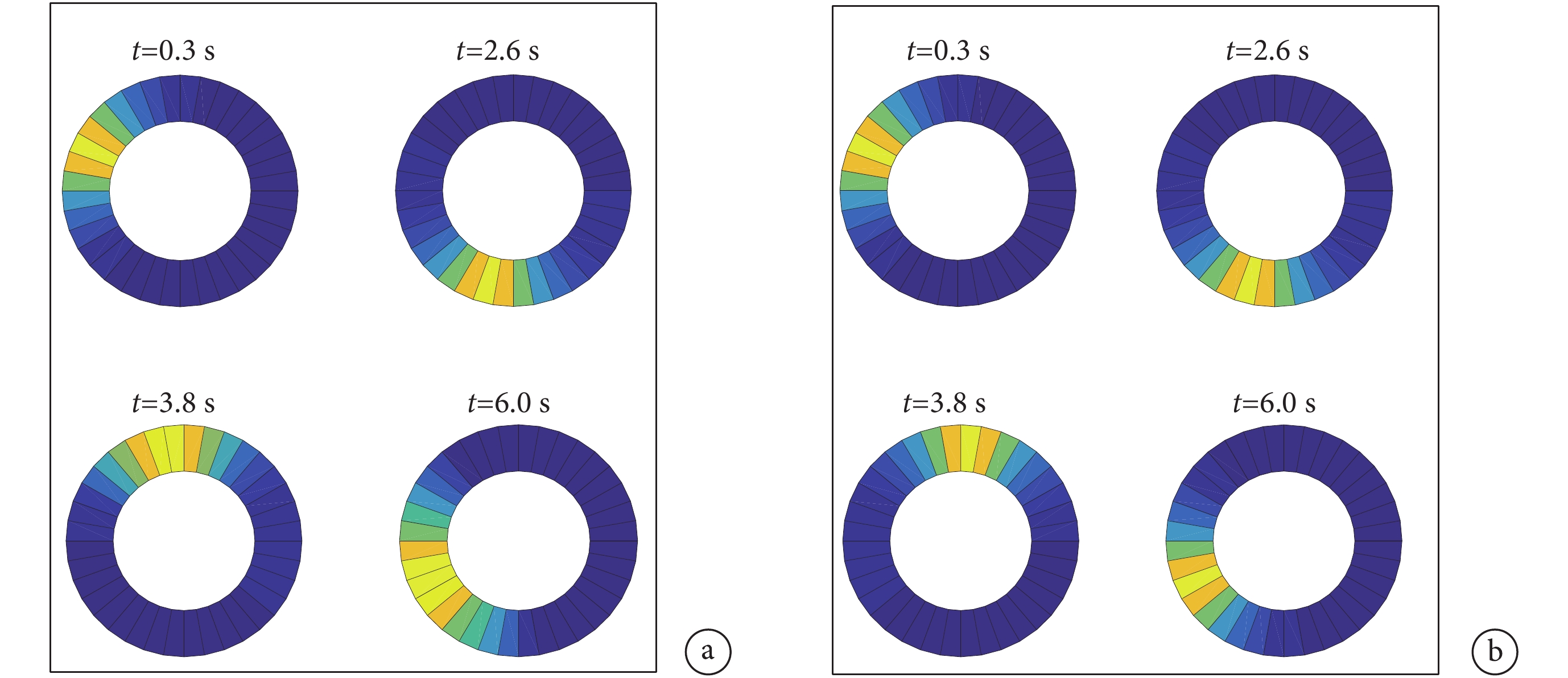 圖12
興奮活動包放電效果圖
圖12
興奮活動包放電效果圖
a.未加入興奮性連接;b.加入興奮性連接
Figure12. Discharge effect diagram of excitatory activity packa. without excitatory connections; b. with excitatory connections
針對興奮活動包擴散的問題,本文在 3.2 節提出了一種頭朝向細胞之間加入興奮性連接的神經網絡模型,通過局部興奮性連接對模型進行調控,加入興奮性連接之后的興奮活動包的放電效果如圖 12b 所示。對比圖 12a 可以看出,改進后的模型中隨著興奮活動包在環狀模型中移動,興奮活動包未出現擴散的情況,表明改進后的頭朝向細胞模型可有效且準確地實現對頭朝向角度的跟隨。
5.3 網格野計算模型實驗驗證
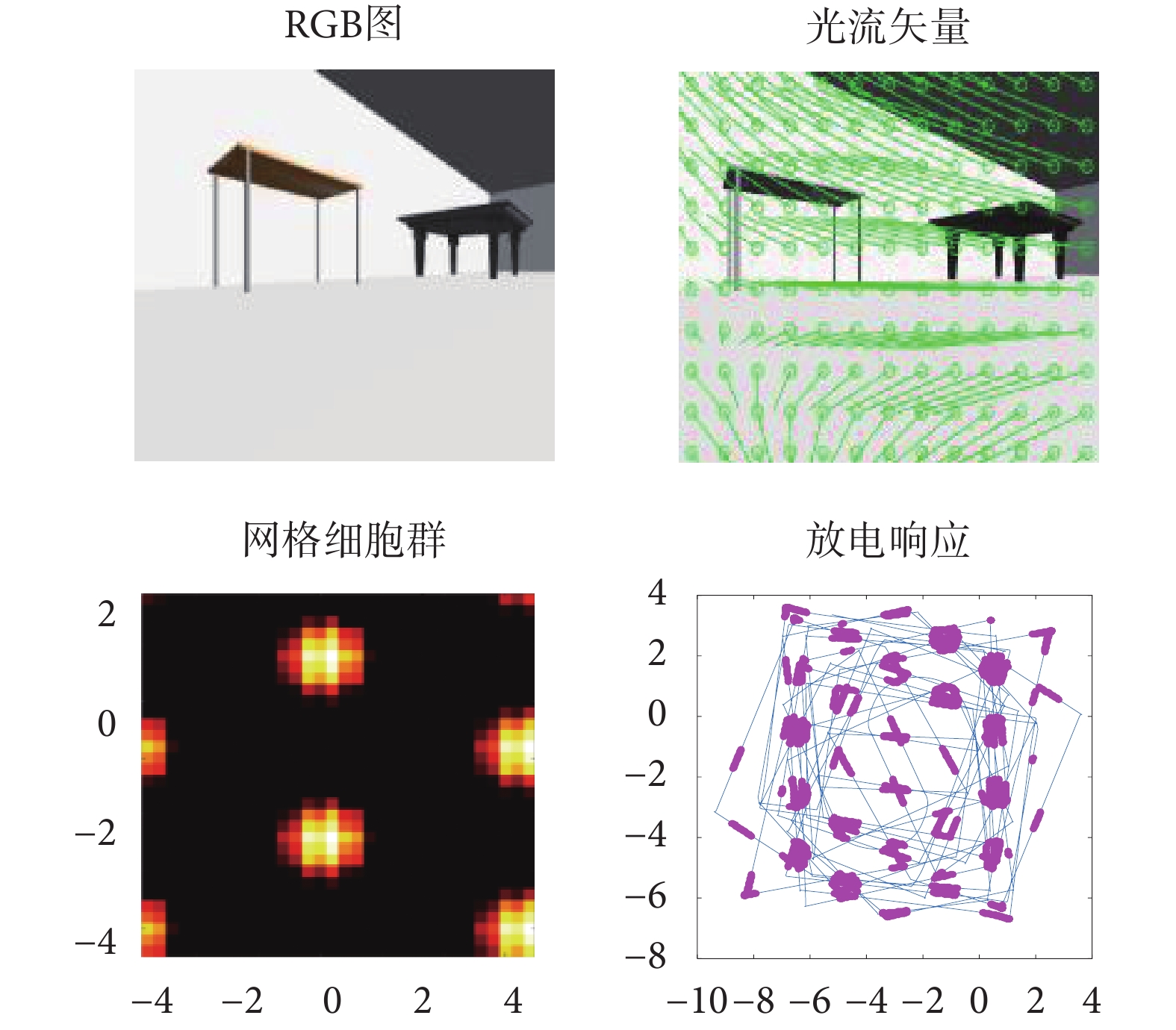
機器人在仿真環境中隨機探索 30 min 得到的仿真結果如圖 13 所示。其中,第一行第一列為機器人采集得到的原始圖像,第二列為計算得到的光流場圖。第二行為網格細胞放電活動圖及當前探索軌跡下得到的網格細胞放電野圖。機器人探索軌跡下得到的網格細胞放電野圖對 15 m*15 m 仿真環境進行了同等比例的映射。圖中藍色線為機器人在仿真環境中探索得到的軌跡線,紫色點代表網格細胞在該位置點的放電活動。實驗結果表明基于感知速度與感知角度的網格細胞模型可實現在特定空間位置上發生穩定的重復性規律放電,得到符合生理學依據的網格細胞周期性六邊形網格野形狀。
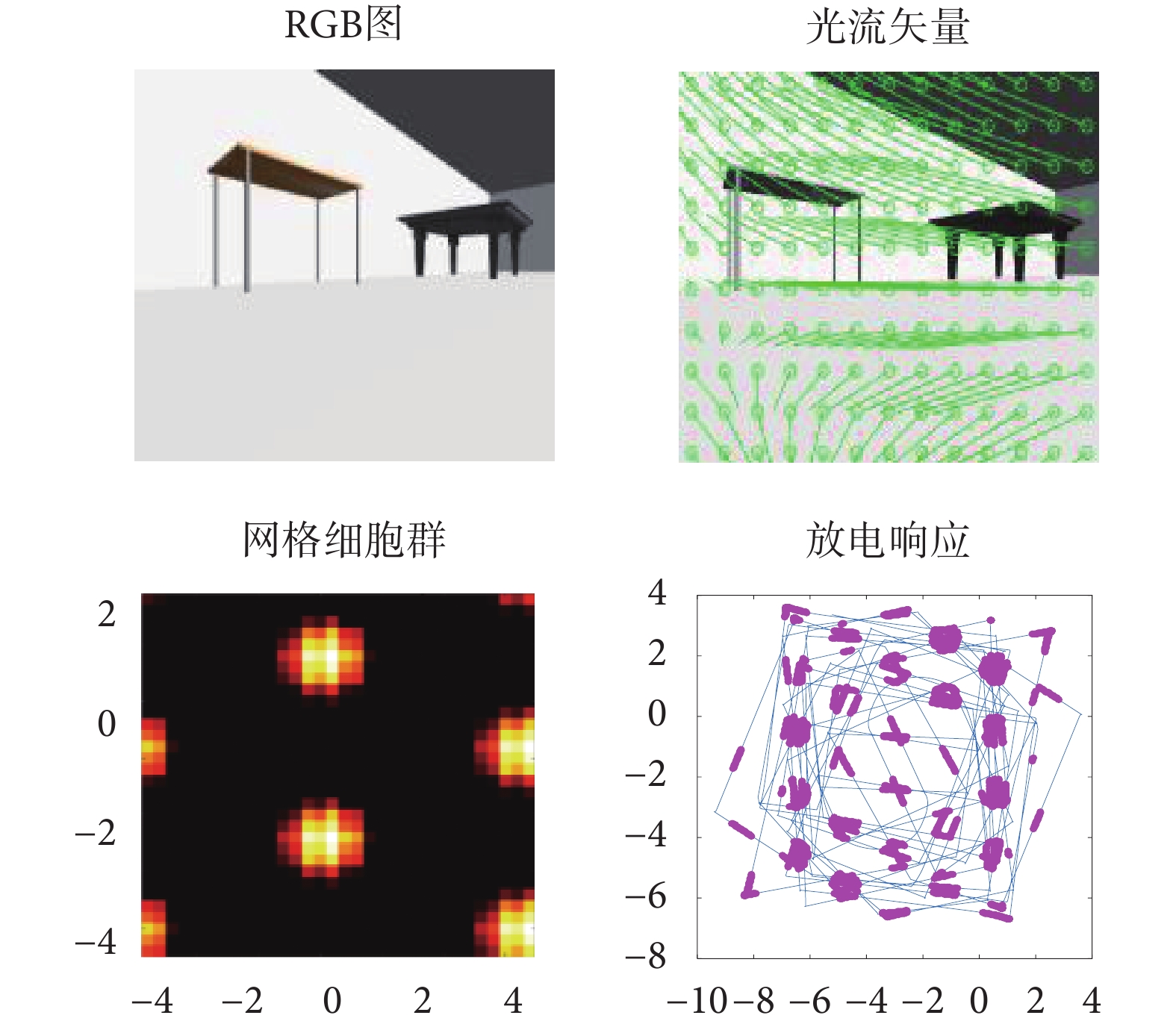 圖13
本文算法運行實驗結果
Figure13.
Experimental results of the algorithm running in this paper
圖13
本文算法運行實驗結果
Figure13.
Experimental results of the algorithm running in this paper
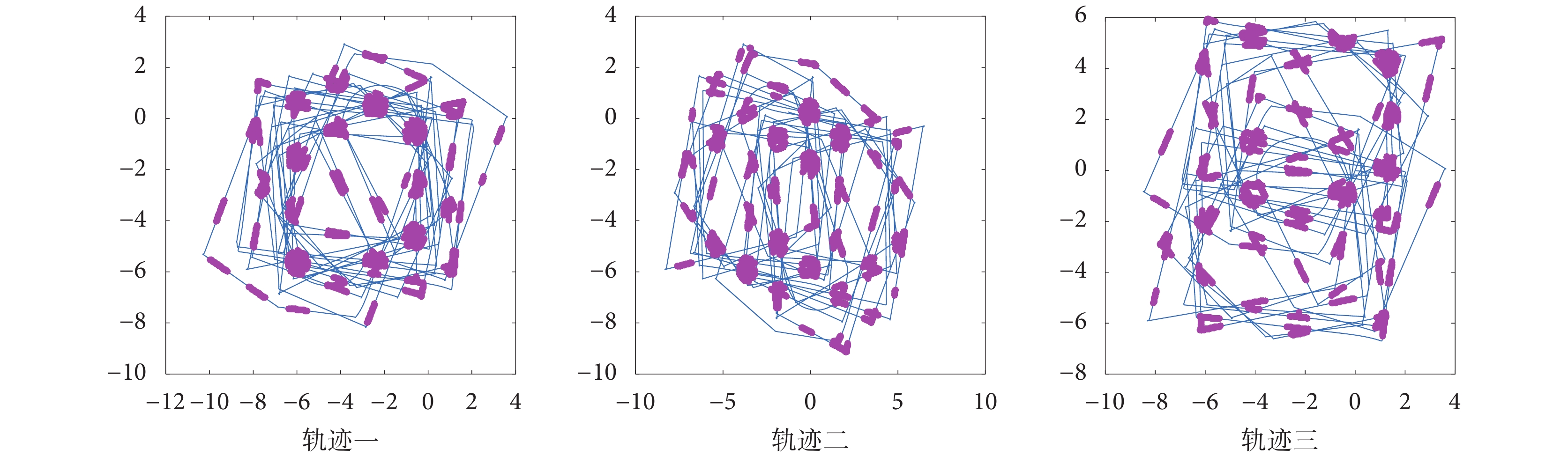
控制機器人在仿真環境中隨機探索三次,分別設定三組仿真實驗中速度細胞、頭朝向細胞的放電率參數以及網格細胞網格野間距相同,得到不同探索軌跡下的網格細胞放電野如圖 14 所示。從圖 14 可以看出不同的探索軌跡下,由感知速度和感知角度所驅動的網格細胞在機器人探索環境的過程中周期性放電并形成了符合生理學實際的六邊形放電野,所形成的網格細胞網格野可以覆蓋到機器人探索的大部分環境,證明了本文所提模型的合理性和有效性。
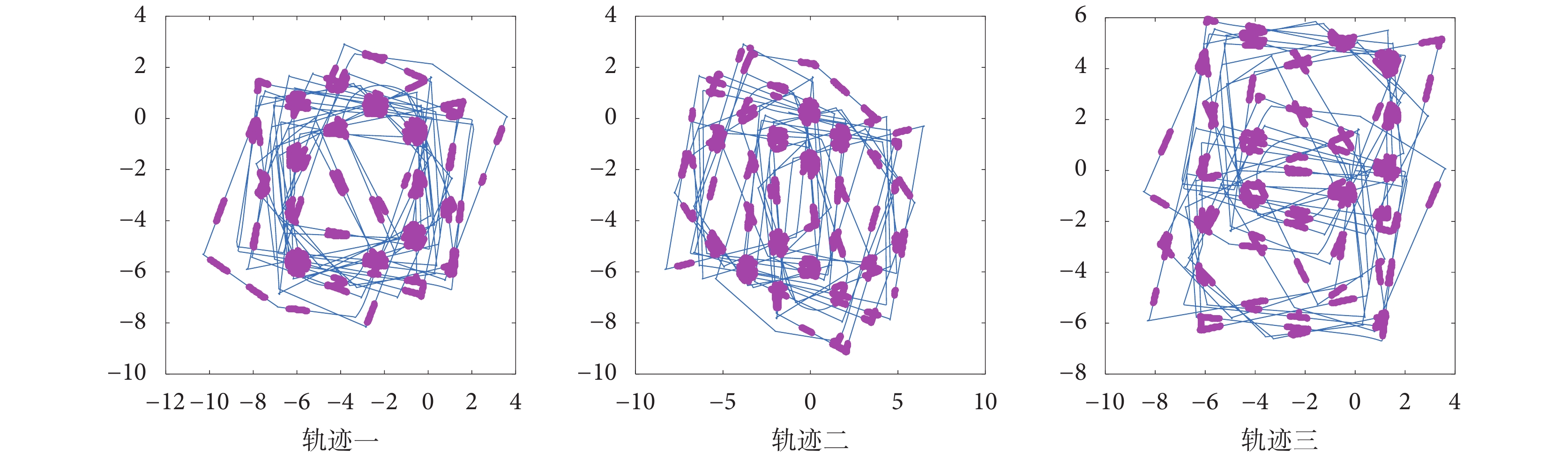 圖14
不同探索軌跡下網格細胞放電響應
Figure14.
Grid cells firing response under different exploration trajectories
圖14
不同探索軌跡下網格細胞放電響應
Figure14.
Grid cells firing response under different exploration trajectories
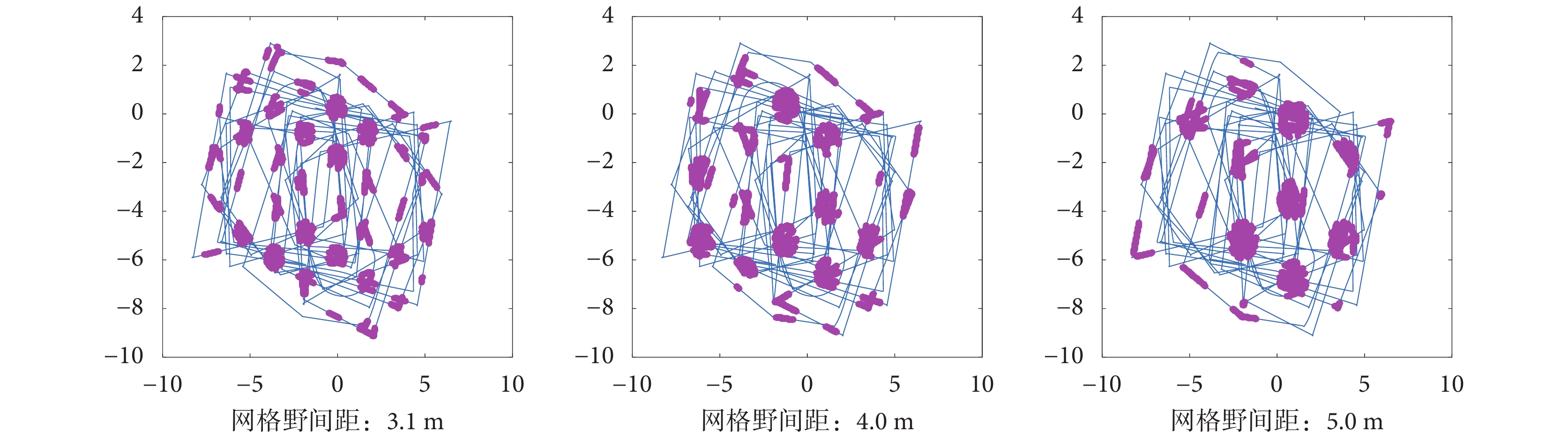
設定頭朝向細胞的放電率參數一致,改變感知速度的輸入模式,得到如圖 15 所示的相同軌跡下網格野間距分別為 3.1、4.0、5.0 m 的網格細胞放電野。可以看出本文模型可以構建出多尺度的網格細胞網格野,符合生理學實際。
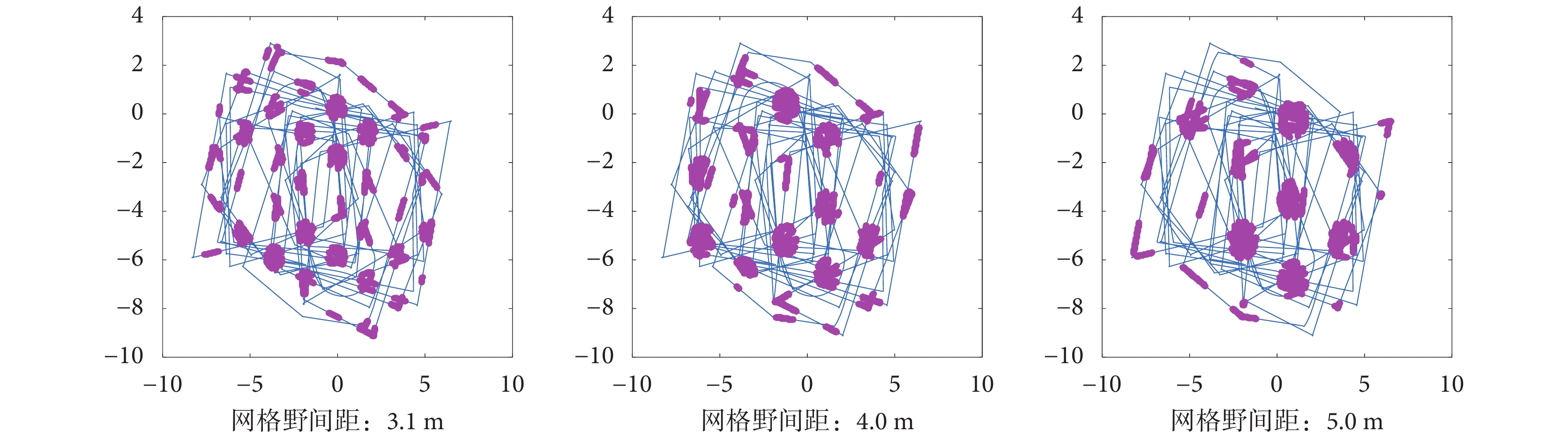 圖15
不同尺度網格細胞放電響應
Figure15.
Grid cells firing response with different scales
圖15
不同尺度網格細胞放電響應
Figure15.
Grid cells firing response with different scales
5.4 網格野計算模型魯棒性測試對比實驗
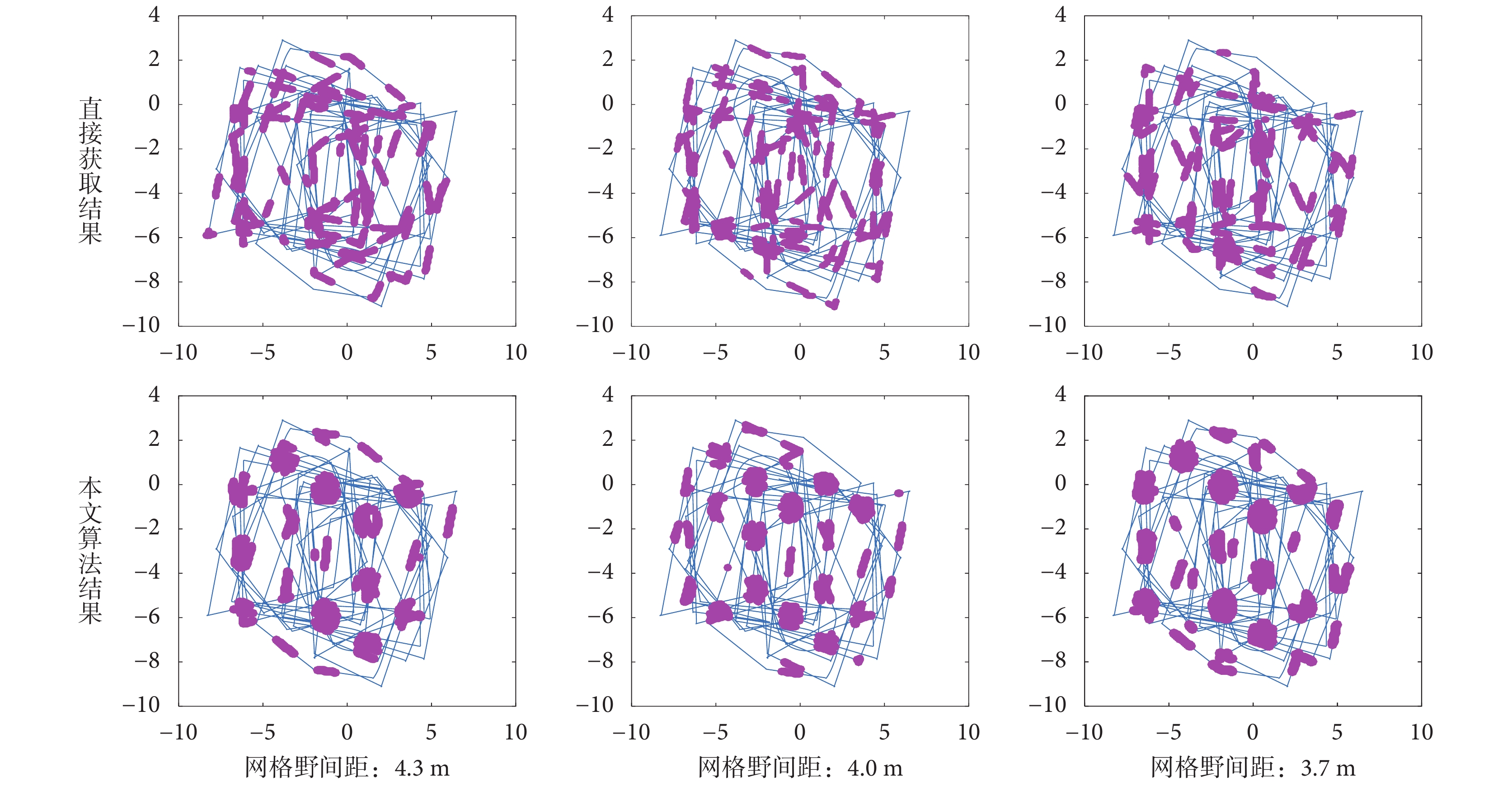
為了驗證本文所構建的計算模型相比于直接由編碼器信息驅動的網格細胞模型具有更強的抗干擾性,做了如下的魯棒性測試對比實驗。為模擬真實環境中由編碼器信息解算的速度存在輕微偏差的現象,人為地對仿真環境中機器人運動的真實速度加入微小的動態干擾,得到如圖 16 所示三組相同軌跡下的不同網格野間距的網格細胞的放電活動圖。
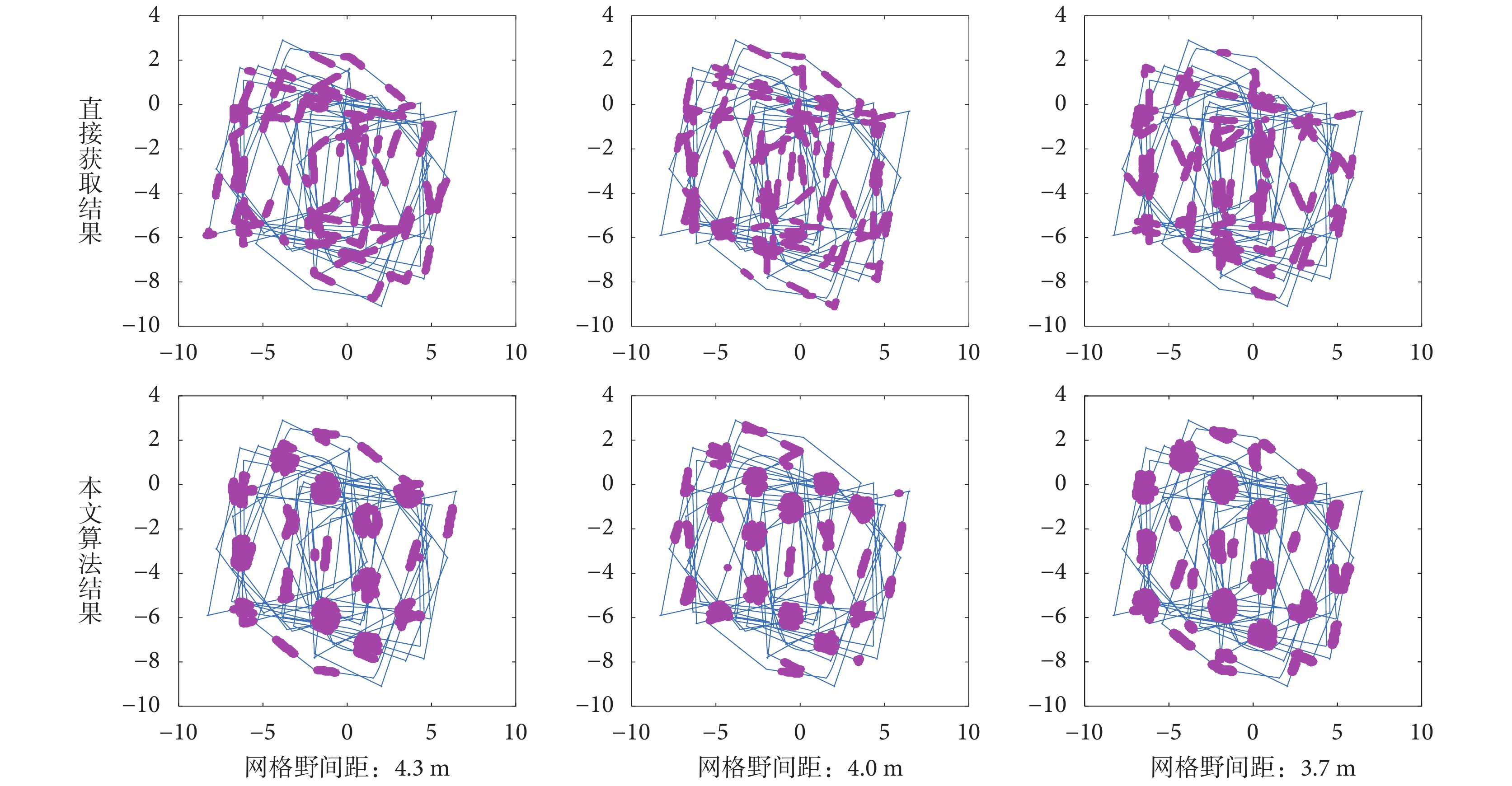 圖16
加入干擾后的網格野計算模型魯棒性測試對比
Figure16.
Robust test of grid field calculation model after adding interference
圖16
加入干擾后的網格野計算模型魯棒性測試對比
Figure16.
Robust test of grid field calculation model after adding interference
可以看出當速度信息加入輕微干擾后,直接由速度和角度驅動的網格細胞的放電野變得散亂無序,不能呈現出周期性的六邊形放電野形狀,而由本文算法得到的網格野依然呈現出穩定的周期性六邊形形狀,網格細胞可以有規律地產生對探索空間的放電活動。傳統方法出現失效,其主要原因是網格細胞路徑積分中所產生的誤差具有累積效應,雖然加入的誤差是微小的,但隨著探索路徑的增長所累加的誤差值也越來越大,最終導致網格細胞放電野出現散亂狀態。而本文所采用的感知速度是融合了視覺速度和本體速度,當編碼器所提供的速度值出現輕微偏差時依靠視覺流獲取的視覺速度可以對其進行糾正,抵消了誤差的累積效應,從而可以保證感知速度不出現嚴重偏差。因此證明了本文所提方法在具有仿生性的同時還具有良好的魯棒性。
5.5 真實環境路徑積分對比實驗
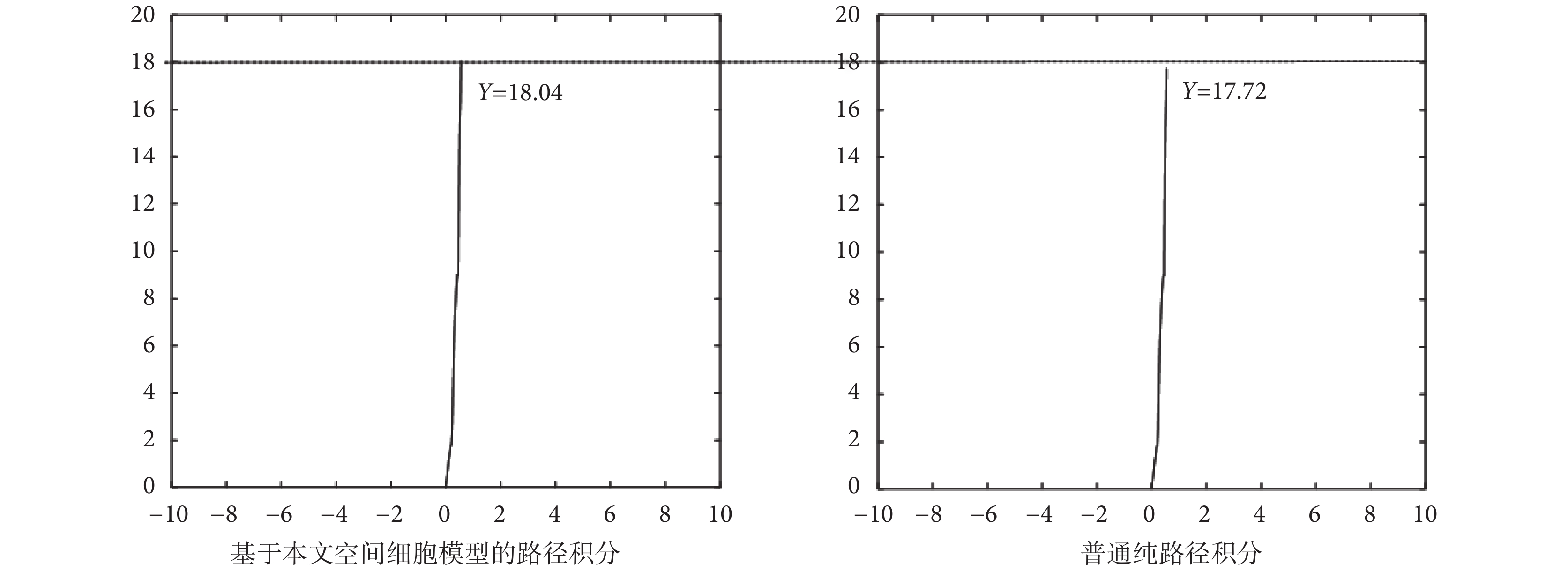
為了證明由感知速度和感知角度驅動的網格細胞路徑積分功能在實際應用中的準確性,本文設計了如下的路徑積分對比實驗。設計實驗在長度為 18 m 的樓道中,由機器人編碼器采集當前的位姿信息,分別進行純路徑積分實驗和依靠空間細胞模型的路徑積分實驗。實驗過程中機器人的運行速度手動控制且最大速度設置為 1 m/s,實驗結果如圖 17 所示,圖中實線為機器人的實際運動軌跡。
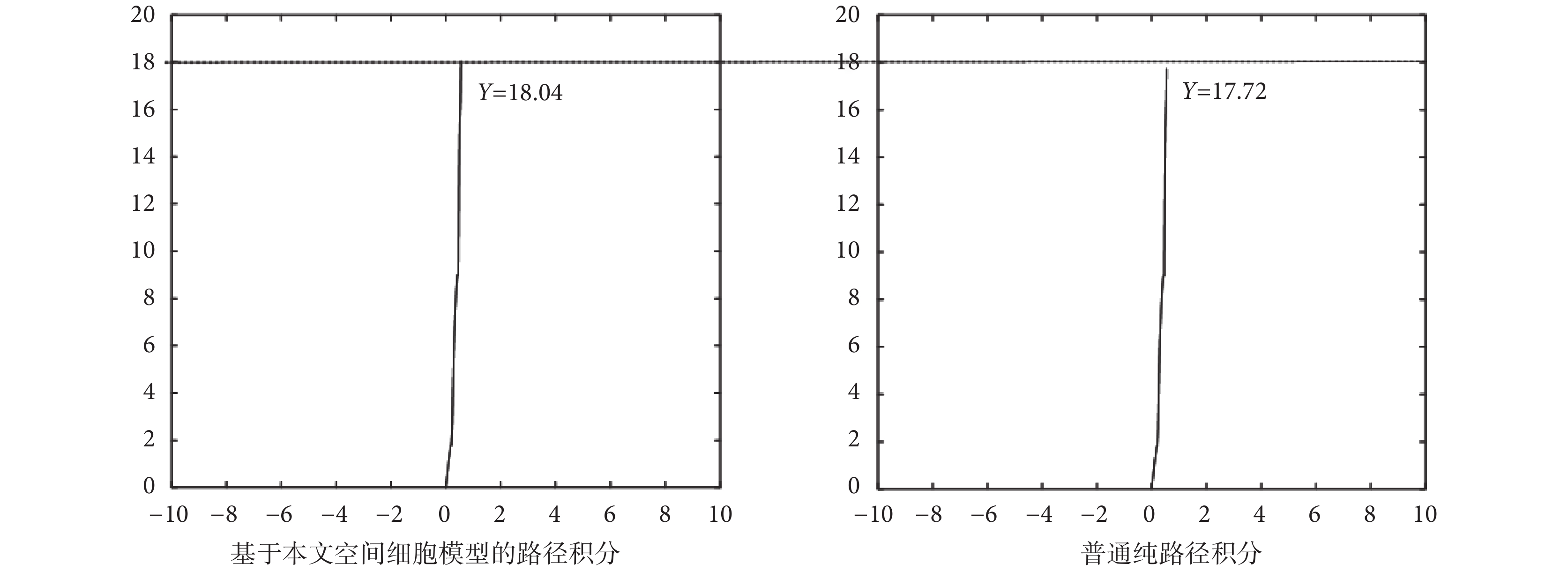 圖17
真實環境路徑積分結果對比圖
Figure17.
Comparison chart of path integration results under real environment
圖17
真實環境路徑積分結果對比圖
Figure17.
Comparison chart of path integration results under real environment
實驗可得,基于本文空間細胞模型的路徑積分結果為 18.04 m,純路徑積分方法得到的結果為 17.72 m,實際樓道長度為 18 m。數據條件不做改變的情況下,純路徑積分的精度要比基于空間細胞模型的路徑積分的精度低。其主要原因是基于感知速度和感知角度的網格細胞模型在路徑積分過程中對數據具有一定的容錯能力,尤其是視覺速度的引入也在一定程度上彌補了硬件設備的誤差以及路面不平的影響。但本文所提方法也不能完全消除累計誤差和積分漂移帶來的影響。
6 結論
本文依照海馬空間細胞的生理學認知機制,采用統一的計算機制對海馬結構中頭朝向細胞、速度細胞與網格細胞這三種空間細胞進行了建模研究。利用加入興奮性連接的一維環狀模型模擬頭朝向細胞的放電機制,以仿生的方式獲取感知角度信息。結合視覺信息與速度細胞模型,提出了一種融合視覺速度和本體速度的感知速度求解方法。利用感知角度信息與感知速度信息驅動網格細胞的二維連續吸引子模型,實現了網格細胞的路徑積分功能,可以得到在探索環境中網格細胞穩定的周期性六邊形網格野形狀。通過仿真實驗驗證了速度細胞模型和頭朝向細胞模型的可靠性。采集仿真環境下的數據集對感知速度模型和感知角度模型進行實驗驗證,驗證了模型的合理性。最后對網格細胞網格野計算模型進行了仿真實驗驗證,在空間探索過程中網格細胞可以生成符合生理學實際的穩定的周期性六邊形放電野,表明了本文算法的合理性。進一步做了魯棒性測試對比實驗,驗證了模型的可靠性,并通過走廊路徑積分對比實驗驗證了所提模型在路徑積分應用中的準確性。本文所提基于感知速度和感知角度的網格野計算模型完全依據海馬生理學認知機制,為進一步研究仿生機器人認知地圖構建與定位導航方法奠定了基礎。
利益沖突聲明:本文全體作者均聲明不存在利益沖突。
引言
生理學研究表明,海馬結構是大鼠進行環境認知的關鍵腦區[1]。海馬結構中存在多種空間細胞,如頭朝向細胞、速度細胞、網格細胞和位置細胞等。1984 年 Ranck 等在大鼠的前下托發現了對頭部方向具有強烈感知作用的神經元細胞,該類神經元細胞的放電率會隨著大鼠頭部朝向的改變而變化,因此被命名為頭朝向細胞[2-3]。1998 年,O’Keefe 等對速度細胞的存在提出了猜想,并認為速度細胞的放電率和大鼠的運動速度呈正相關關系。2015 年 Kropff 等[4]通過實驗證實了速度細胞的存在,并引起了學術界對空間認知過程中速度信號的編碼規則與作用機制的思考。研究表明,通過整合速度細胞提供的信息,大鼠可以實現對運動速度的感知,這為大鼠構建精確的認知地圖提供了生理學依據。2005 年 Hafting 等[5]通過變換實驗箱大小和形狀的實驗,發現了對空間位置具有強烈放電作用的網格細胞,網格細胞會在大鼠所探索的空間中發生穩定的周期性放電,并形成六邊形的網格野覆蓋大鼠遍歷的整個空間環境[6-7]。網格野可以起到度量大鼠所遍歷空間的距離和角度的作用,從而指導大鼠確定空間坐標,實現自我定位,完成對空間環境的認知。振蕩干涉模型[8-11]和吸引子模型[12-13]是兩種經典的網格細胞計算模型。
行為學研究表明,動物能夠利用自運動線索保持相對環境重要位置的矢量關系,并將空間中的各個位點的信息進行計算和整合,使這種矢量關系不斷得到更新,這一過程被稱為路徑積分[14-15]。研究表明,網格細胞在大鼠大腦中充當路徑整合的角色[16]。速度和角度是路徑積分過程中的主要輸入信息,并且該過程是網格細胞網格野形成的主要機制。其中,頭朝向細胞提供感知方向信息[17],速度細胞提供不含方向信息的運動速度信號。然而,在大鼠腦內直接參與環境認知過程的速度信息并非僅由速度細胞提供,而是由多種信號共同編碼得到的一種感知速度。直接參與路徑積分過程的感知速度是由包括視覺因素在內的多種因素影響而產生的。波士頓大學 Raudies 等[18]的改變視覺-空間輸入的實驗以及清華大學謝康寧[19]的控制大鼠運動的實驗都證明了上述結論。
先前的網格細胞網格野計算模型,直接使用由編碼器解算的速度值驅動網格細胞吸引子,沒有對速度細胞進行建模研究,這種方法缺乏生理學依據,并且單純依靠編碼器信息驅動網格細胞吸引子的方法抗干擾能力相對較差[13]。2018 年本團隊[14]建立了一種頭朝向細胞計算模型,該計算模型給定了頭朝向細胞間的調整內核,但未對細胞之間的權重連接給出分析,模型不夠完善且對頭朝向信號的跟隨不夠精確。針對以上問題,本文根據海馬結構中的空間細胞的生理學研究[20]以及感知速度的獲取模式研究,首先,對速度細胞進行建模并且提出了一種融合視覺速度和本體速度從而獲得感知速度的計算方法;其次,利用加入興奮性連接的一維環狀細胞模型模擬了頭朝向細胞的放電機制,提出了一種感知角度的獲取方法;最后,利用感知速度和感知角度驅動網格細胞吸引子模型,實現網格細胞對空間環境的編碼。
1 模型架構
本文所構建的基于感知速度與感知角度的網格野計算模型如圖 1 所示。首先,攝像頭采集環境的圖像信息,編碼器采集機器人的速度信息與方向信息。然后,融合視覺速度和本體速度以仿生的方式獲取感知速度信息。接著,將頭朝向細胞建模成加入興奮性連接的一維環狀模型,以仿生的方式獲取感知角度信息。最后,將感知速度信息和感知角度信息輸入網格細胞二維連續吸引子模型,驅動網格細胞板上興奮活動包的移動,實現網格細胞對空間環境的編碼以及路徑積分功能。
圖1
基于感知速度與感知角度的網格野計算模型
Figure1.
Grid field calculation model based on perceived speed and perceived angle
2 感知速度的獲取
感知速度是由多種信號協同編碼產生的,且感知速度與實際速度之間不可避免地會存在誤差。影響感知速度的主要因素有:視覺流和本體感覺。視網膜圖像的連續變化稱為視覺流,這種連續變化會引起生物體對自身速度的估計。本體感覺指的是由速度細胞構成的速度碼系統,該系統獲取不含方向信息的運動速率作為本體速度[21]。基于此,本文提出一種基于速度細胞和視覺信息的感知速度求解方法。圖 2 為感知速度獲取原理示意圖。
圖2
感知速度獲取原理示意圖
Figure2.
Schematic diagram of the principle of perceived speed acquisition
2.1 基于光流法的視覺速度獲取
將光流算法應用到本模型中,采用經典的 Farneback 光流算法[22]計算連續圖像間的光流場,關于光流算法的具體內容本文不再進行闡述。根據亮度恒定假設、圖像梯度恒定假設以及空間一致性計算所有圖像序列的光流矢量,光流矢量關于圖像豎直方向分量大小的絕對值記為。
|
采用粒子群優化的徑向基函數(radial basis function,RBF)神經網絡[23-24]作為通過視覺估計線速度的算法。為避免陷入局部最小值,采用粒子群算法對神經網絡權值進行優化,取光流矢量關于圖像豎直方向分量大小的絕對值作為神經網絡的輸入樣本,以每一幀圖像拍攝時機器人實際運行的線速度作為輸出樣本,對神經網絡進行訓練。權值優化算法流程圖如圖 3 所示。
圖3
粒子群優化 RBF 神經網絡權值算法流程圖
Figure3.
Algorithm flow chart of particle swarm optimization of the weight of RBF neural network
2.2 基于速度細胞模型的本體速度獲取
生理學研究表明,速度細胞的放電率與大鼠的運動速度呈正相關關系,并且當大鼠處于靜止狀態時速度細胞也會產生放電現象[4, 25]。基于生理學實際,本文構建了如下的速度細胞模型。
定義速度細胞的放電率公式為
|
|
|
式(2)中 代表第 個速度細胞的放電率,、 大于零,其中, 為速度細胞感知速度的基礎偏置量, 為速度細胞對速度值的感知比例。 小于零,保證了速度細胞對運動速度具有良好的跟隨性。如式(3)所示, 為增加了擾動之后速度細胞所感知到的速度信息。 為速度細胞感知速度的擾動值,如式(4)所示擾動信號服從標準正態分布。
為使本文所構建的速度細胞模型更符合生理學依據,定義速度細胞置信度為大鼠的運動速度和放電率之間的 Pearson 相關系數。
|
式(5)中 為所有速度細胞放電率的平均值, 為運動速度的平均值, 為速度細胞群中速度細胞的數量。公式計算所得 為速度細胞的置信度,閾值范圍為 0.94~0.97,當 小于閾值時檢查速度細胞模型,并從中選擇離群值淘汰,得到精確度更高的速度細胞群,從而使得歸一化后計算的速度細胞放電率的數學期望值更具可靠性。對滿足閾值條件的速度細胞群做如下處理。
首先,計算單個速度細胞的放電率在速度細胞群放電率中的權值 。
|
式(6)中 為剔除離群值之后速度細胞的數量,然后對所有速度細胞的放電率做歸一化處理。
|
式(7)中 為歸一化處理后第 個速度細胞的放電率, 為速度細胞群中速度細胞放電率的標準差, 的計算公式如式(8)所示。
|
最后根據式(9)求解速度細胞群放電率的數學期望 ,即所有速度細胞放電率的加權平均值。
|
2.3 感知速度的求解
當視覺流的連續變化受到附加速度的影響較大時,會由于視覺對比度產生對自身速度的嚴重錯估。當編碼器信息受到路面環境的輕微擾動時,會使得對本體速度的估計產生偏差。因此在求解感知速度 的時候,采用公式(10)對感知速度進行求解。
|
式(10)中, 和 分別為速度細胞放電率的數學期望 和神經網絡的輸出 轉化至同一量綱下的結果, 代表權值系數, 代表調整閾值,當 和 之間的差值大于 時,判斷視覺對比度引起了對自身速度的嚴重錯估,令感知速度為 ;當 和 之間的差值小于等于 時,判斷視覺速度估算正確,同時為了減小由于擾動導致的本體速度估值的偏差,令 和 通過權值系數 求解得到的加權平均值作為感知速度 。
3 感知角度的獲取
圖 4 為感知角度獲取原理示意圖,通過一維環狀細胞模型對頭朝向細胞進行建模,獲取環狀模型上興奮活動包的位置實現感知角度的獲取。
圖4
感知角度獲取原理示意圖
Figure4.
Schematic diagram of the principle of perceived angle acquisition
3.1 頭朝向細胞的建模
頭朝向細胞具有如下放電特性[26-27]:以水平方向為起點,細胞的放電強度隨著大鼠頭朝向角度的增加而增加,當大鼠的頭朝向到達該頭朝向細胞的最佳頭部方向時,放電率達到峰值,然后跟隨頭朝向角度的增大而逐漸偏離最佳方向,并且放電強度也降低[14]。根據頭朝向細胞的生理學研究事實,定義頭朝向細胞的放電率公式如式(11)所示。
|
式(11)中, 代表頭朝向細胞 的放電率, 代表當前頭朝向角度, 代表該頭朝向細胞的最優方向,K 決定放電率曲線的銳度, 和 分別決定頭朝向細胞的放電基準值和放電率峰值大小。根據公式(11),設定 等于 0、 等于 0.01 以及 K 等于 4.53 的優先方向為 100° 的頭朝向細胞的放電特性如圖 5 所示。
圖5
頭朝向細胞放電特性示意圖
Figure5.
Schematic diagram of discharge characteristics of head direction cell
3.2 一維環狀頭朝向細胞的建模
將順序排列的頭朝向細胞首尾連接形成一個封閉的環形,第 個頭朝向細胞對應的最優方向為 ,并且可以表示為相對主朝向 的角偏移量, 的計算公式如式(12)所示。
|
其中 代表頭朝向細胞的編號, 代表一維環狀模型中頭朝向細胞的個數。
根據頭朝向細胞的放電特性,可知在環狀細胞模型中將會出現一個高斯帽狀的興奮活動包,隨著頭朝向角度的變化,興奮活動包會在環狀細胞模型上移動。獲取興奮活動包在環狀模型上的位置,即可得知當前的頭朝向信息。
隨著興奮活動包在環狀細胞模型上長時間移動,興奮活動包會逐漸擴散,將導致從環狀細胞模型上獲取的角度信息與真實角度信息產生誤差。因此,本文提出一種頭朝向細胞之間加入興奮連接的神經網絡模型,頭朝向細胞之間的連接強度計算公式如式(13)所示[26]。
|
其中,、 為常系數, 是興奮性連接基準值, 是興奮參數權值。 是常系數,、 分別表示頭朝向細胞 和頭朝向細胞 在環狀模型的位置坐標,從公式(13)可以看出頭朝向細胞之間的興奮性連接強度只和相互連接的頭朝向細胞之間的角度差有關。
由興奮性連接產生的頭朝向細胞的放電變化量為
|
式(14)中, 代表頭朝向細胞之間的興奮性連接權值, 代表頭朝向細胞 的放電率, 代表一維環狀細胞模型中頭朝向細胞的個數。興奮性傳遞之后,將所有頭朝向細胞的放電率與 0 進行比較,以保證所有頭朝向細胞的放電率都不小于零,然后做歸一化處理,其數學表達式如式(15)和(16)所示。
|
|
4 網格細胞網格野計算模型
在沒有任何外界輸入信息的環境中,網格細胞能夠根據自運動線索中的感知速度和感知角度進行精確的路徑積分,網格細胞的放電活動被認為能夠形成空間環境的度量地圖[28]。本文在 Burak 等[13]提出的網格細胞模型基礎上進行了改進,提出了一種基于感知速度與感知角度的網格野計算模型,本文模型中網格細胞接收的輸入信息不再是未經處理的編碼器信息,而是將編碼器信息以前面所提仿生的方式進行處理,獲得更具有生理學依據且抗干擾性更強的感知速度和感知角度。網格細胞網格野計算模型結構圖如圖 6 所示。
圖6
網格細胞網格野計算模型結構圖
Figure6.
Structure diagram of grid field calculation model
網格細胞吸引子受兩方面的驅動:一是網格細胞之間的抑制性遞歸輸入;二是來自上游皮層的興奮性投射信息。使用二維連續吸引子模型對網格細胞進行建模,每個神經元對應一個吸引子[14]。隨著大鼠在空間環境中的不斷探索,遍歷的范圍也逐漸增大,然而表示空間相對位置的神經元不能無限增加,從而會涉及到吸引子網絡的邊界問題,所以將神經面板的上下邊界和左右邊界相互連接,形成二維環狀吸引子模型[29]。二維的神經面板中網格細胞的神經元動力學等式為
|
其中, 是神經元相應的時間常量,傳遞函數 是一個非線性整流函數,當 時 ,當 時 , 時刻當前位置神經元 的狀態為 , 是神經面板上神經元 到神經元 的連接權值, 是神經面板上周圍神經元到神經元 的抑制性遞歸輸入,是來自上游皮層的興奮性投射信息。假定神經面板的長度為 ,那么整個神經網絡有 個神經元,每個神經元 都有一個優先朝向,并且優先朝向信息由頭朝向細胞產生的感知角度確定。感知角度信息確定輸出權值的改變方向,網格細胞的遞歸連接權值為
|
|
權值矩陣的分布將形成中間高兩邊低的墨西哥帽狀形狀,其中間位置為 ,為對 方向的感知速度進行積分之后的周期性重置距離。在所有的實驗中設定 ,,其中 為神經面板上網格的周期。
下面給出感知速度與感知角度對網格細胞吸引子的作用機制。定義 時刻大鼠的頭朝向角度為 ,沿著該方向上的感知速度為 ,那么沿著方向 的感知速度為
|
對速度進行積分得到 方向的位移為
|
分別定義放電周期 和相位 來決定感知速度的輸入模式,其中 決定網絡速度響應的增益,放電周期 為一固定值,那么對 方向的感知速度進行積分之后的周期性重置距離為
|
定義網格細胞所接收的興奮性輸入為
|
|
代表了對輸入信息的耦合并通過感知角度信息確定了網格細胞吸引子的移動方向。
5 實驗驗證
利用仿真實驗和物理實驗對本文所構建的模型進行驗證。為驗證所構建的感知角度和感知速度模型的合理性以及網格野計算模型的有效性和魯棒性,采用 ROS 自帶的 Gazeboard 仿真器搭建一個 15 m*15 m 的虛擬環境并控制機器人對該環境進行探索,采集里程計信息及環境 RGB 圖像進行實驗驗證。通過實際機器人平臺進行了走廊環境路徑積分實驗,驗證了由感知速度和感知角度驅動的網格細胞路徑積分功能的精確性。仿真實驗環境如圖 7 所示。
圖7
仿真實驗界面
Figure7.
Simulation experiment interface
5.1 感知速度模型實驗驗證
利用機器人平臺在虛擬環境下探索 30 min,實時獲取機器人移動過程中的里程計信息及 RGB 圖像,計算得到每幀圖像拍攝時機器人的移動速度及連續圖像間的光流矢量。采用 Farneback 光流算法計算得到的虛擬環境下的光流場效果如圖 8 所示。設計 RBF 神經網絡作為通過視覺估計線速度的算法,取光流矢量關于圖像豎直方向分量大小的絕對值作為輸入信息,以每一幀圖像拍攝時機器人實際運行的線速度作為輸出樣本,并采用粒子群算法對神經網絡權值以及 RBF 的中心和寬度進行優化。設置粒子群迭代次數為 250 次,種群規模為 30,粒子維度 12,加速度因子 。設定 Farneback 光流算法計算光流場的分辨率為 10*130,即 RBF 神經網絡有 130 個輸入節點,1 個輸出節點,通過正交最小二乘法計算得到隱含層中心數為 22。設定神經網絡學習率為 0.05,均方誤差為 0.000 3,迭代次數為 240 次,以上參數的設定值為多次實驗后得到的經驗值。
圖8
仿真環境下的圖像光流場效果圖
Figure8.
Optical flow vector of image in simulation environment
對 RBF 神經網絡訓練的過程是一個參數尋優的過程,將粒子群中的每一個體分量映射到網絡中的權值,對每一個體對應的神經網絡輸入機器人采集的連續圖像幀并進行訓練,網絡中權值的優化過程是一個反復迭代的過程,迭代的終止條件是最大迭代次數。對訓練后的網絡進行測試,由 RBF 神經網絡得出的視覺速度與機器人的實際運動速度對比圖如圖 9 所示。
圖9
視覺速度和真實速度對比圖
Figure9.
Comparison chart of visual speed and real speed
對 2.2 節所提的速度細胞數學模型進行實驗驗證,為保證模型的合理性與準確性,對速度細胞放電率參數設定如下:、、、 均設定為服從正態分布,其中, 服從均值為 4、方差為 2 的正態分布; 服從均值為 1.5、方差為 3 的正態分布; 服從均值為-0.005、方差為 0.001 的正態分布;Vr 服從均值為 0、方差為 1 的正態分布。同時,設定速度細胞個數為 20 個。對速度細胞數學模型進行實驗驗證,得到如圖 10 所示的不同參數值下的速度細胞放電率對比圖。圖中所示的速度細胞的放電率與運動速度之間的相關系數均大于規定的閾值,表明了本文模型所構建的速度細胞的可靠性,且符合速度細胞放電率與大鼠的運動速度呈正相關關系這一生理學現象。
圖10
不同參數值下的速度細胞放電率圖
Figure10.
Speed cell firing rate map under different parameter values
為模擬真實環境中依靠編碼器信息解算速度值存在干擾與偏差這一現象,將仿真環境中得到的真實速度加入動態的輕微擾動,模型中令擾動服從標準正態分布。根據 2.2 節所提算法,速度細胞群對加入擾動之后的速度信息進行處理并解碼獲得本體速度。根據本文融合算法,將本體速度與通過 RBF 神經網絡獲得的視覺速度進行加權平均,得到感知速度 ,感知速度與真實速度對比圖如圖 11 所示。
圖11
感知速度和真實速度對比圖
Figure11.
Comparison chart of perceived speed and real speed
從圖 9 和 圖 11 可以看出,基于光流解算得到的視覺速度基本可以實現對當前運動速度的估計,基于視覺速度和本體速度融合得到的感知速度與真實速度基本接近,同時又不完全依賴于機器人本身所攜帶的編碼器對速度的測量。表明所構建的感知速度獲取模型符合直接參與環境認知過程的速度信息是多種信號協同編碼的結果這一生理學實際,且感知速度與實際速度之間不可避免地存在誤差。
5.2 感知角度模型實驗驗證
本文中將頭朝向細胞建模成一維環狀模型,根據頭朝向細胞的放電特性可知在環狀細胞模型上將會產生一個高斯帽狀的興奮活動包。隨著興奮活動包在環狀細胞模型上的長時間移動興奮活動包會逐漸擴散,這會導致從環狀細胞模型上獲取的感知角度信息與真實的角度信息之間產生誤差。對未加入興奮連接的環狀細胞模型進行仿真實驗得到的興奮活動包擴散的效果如圖 12a 所示。
圖12
興奮活動包放電效果圖
a.未加入興奮性連接;b.加入興奮性連接
Figure12. Discharge effect diagram of excitatory activity packa. without excitatory connections; b. with excitatory connections
針對興奮活動包擴散的問題,本文在 3.2 節提出了一種頭朝向細胞之間加入興奮性連接的神經網絡模型,通過局部興奮性連接對模型進行調控,加入興奮性連接之后的興奮活動包的放電效果如圖 12b 所示。對比圖 12a 可以看出,改進后的模型中隨著興奮活動包在環狀模型中移動,興奮活動包未出現擴散的情況,表明改進后的頭朝向細胞模型可有效且準確地實現對頭朝向角度的跟隨。
5.3 網格野計算模型實驗驗證
機器人在仿真環境中隨機探索 30 min 得到的仿真結果如圖 13 所示。其中,第一行第一列為機器人采集得到的原始圖像,第二列為計算得到的光流場圖。第二行為網格細胞放電活動圖及當前探索軌跡下得到的網格細胞放電野圖。機器人探索軌跡下得到的網格細胞放電野圖對 15 m*15 m 仿真環境進行了同等比例的映射。圖中藍色線為機器人在仿真環境中探索得到的軌跡線,紫色點代表網格細胞在該位置點的放電活動。實驗結果表明基于感知速度與感知角度的網格細胞模型可實現在特定空間位置上發生穩定的重復性規律放電,得到符合生理學依據的網格細胞周期性六邊形網格野形狀。
圖13
本文算法運行實驗結果
Figure13.
Experimental results of the algorithm running in this paper
控制機器人在仿真環境中隨機探索三次,分別設定三組仿真實驗中速度細胞、頭朝向細胞的放電率參數以及網格細胞網格野間距相同,得到不同探索軌跡下的網格細胞放電野如圖 14 所示。從圖 14 可以看出不同的探索軌跡下,由感知速度和感知角度所驅動的網格細胞在機器人探索環境的過程中周期性放電并形成了符合生理學實際的六邊形放電野,所形成的網格細胞網格野可以覆蓋到機器人探索的大部分環境,證明了本文所提模型的合理性和有效性。
圖14
不同探索軌跡下網格細胞放電響應
Figure14.
Grid cells firing response under different exploration trajectories
設定頭朝向細胞的放電率參數一致,改變感知速度的輸入模式,得到如圖 15 所示的相同軌跡下網格野間距分別為 3.1、4.0、5.0 m 的網格細胞放電野。可以看出本文模型可以構建出多尺度的網格細胞網格野,符合生理學實際。
圖15
不同尺度網格細胞放電響應
Figure15.
Grid cells firing response with different scales
5.4 網格野計算模型魯棒性測試對比實驗
為了驗證本文所構建的計算模型相比于直接由編碼器信息驅動的網格細胞模型具有更強的抗干擾性,做了如下的魯棒性測試對比實驗。為模擬真實環境中由編碼器信息解算的速度存在輕微偏差的現象,人為地對仿真環境中機器人運動的真實速度加入微小的動態干擾,得到如圖 16 所示三組相同軌跡下的不同網格野間距的網格細胞的放電活動圖。
圖16
加入干擾后的網格野計算模型魯棒性測試對比
Figure16.
Robust test of grid field calculation model after adding interference
可以看出當速度信息加入輕微干擾后,直接由速度和角度驅動的網格細胞的放電野變得散亂無序,不能呈現出周期性的六邊形放電野形狀,而由本文算法得到的網格野依然呈現出穩定的周期性六邊形形狀,網格細胞可以有規律地產生對探索空間的放電活動。傳統方法出現失效,其主要原因是網格細胞路徑積分中所產生的誤差具有累積效應,雖然加入的誤差是微小的,但隨著探索路徑的增長所累加的誤差值也越來越大,最終導致網格細胞放電野出現散亂狀態。而本文所采用的感知速度是融合了視覺速度和本體速度,當編碼器所提供的速度值出現輕微偏差時依靠視覺流獲取的視覺速度可以對其進行糾正,抵消了誤差的累積效應,從而可以保證感知速度不出現嚴重偏差。因此證明了本文所提方法在具有仿生性的同時還具有良好的魯棒性。
5.5 真實環境路徑積分對比實驗
為了證明由感知速度和感知角度驅動的網格細胞路徑積分功能在實際應用中的準確性,本文設計了如下的路徑積分對比實驗。設計實驗在長度為 18 m 的樓道中,由機器人編碼器采集當前的位姿信息,分別進行純路徑積分實驗和依靠空間細胞模型的路徑積分實驗。實驗過程中機器人的運行速度手動控制且最大速度設置為 1 m/s,實驗結果如圖 17 所示,圖中實線為機器人的實際運動軌跡。
圖17
真實環境路徑積分結果對比圖
Figure17.
Comparison chart of path integration results under real environment
實驗可得,基于本文空間細胞模型的路徑積分結果為 18.04 m,純路徑積分方法得到的結果為 17.72 m,實際樓道長度為 18 m。數據條件不做改變的情況下,純路徑積分的精度要比基于空間細胞模型的路徑積分的精度低。其主要原因是基于感知速度和感知角度的網格細胞模型在路徑積分過程中對數據具有一定的容錯能力,尤其是視覺速度的引入也在一定程度上彌補了硬件設備的誤差以及路面不平的影響。但本文所提方法也不能完全消除累計誤差和積分漂移帶來的影響。
6 結論
本文依照海馬空間細胞的生理學認知機制,采用統一的計算機制對海馬結構中頭朝向細胞、速度細胞與網格細胞這三種空間細胞進行了建模研究。利用加入興奮性連接的一維環狀模型模擬頭朝向細胞的放電機制,以仿生的方式獲取感知角度信息。結合視覺信息與速度細胞模型,提出了一種融合視覺速度和本體速度的感知速度求解方法。利用感知角度信息與感知速度信息驅動網格細胞的二維連續吸引子模型,實現了網格細胞的路徑積分功能,可以得到在探索環境中網格細胞穩定的周期性六邊形網格野形狀。通過仿真實驗驗證了速度細胞模型和頭朝向細胞模型的可靠性。采集仿真環境下的數據集對感知速度模型和感知角度模型進行實驗驗證,驗證了模型的合理性。最后對網格細胞網格野計算模型進行了仿真實驗驗證,在空間探索過程中網格細胞可以生成符合生理學實際的穩定的周期性六邊形放電野,表明了本文算法的合理性。進一步做了魯棒性測試對比實驗,驗證了模型的可靠性,并通過走廊路徑積分對比實驗驗證了所提模型在路徑積分應用中的準確性。本文所提基于感知速度和感知角度的網格野計算模型完全依據海馬生理學認知機制,為進一步研究仿生機器人認知地圖構建與定位導航方法奠定了基礎。
利益沖突聲明:本文全體作者均聲明不存在利益沖突。