具有位置調諧特性的位置細胞對于大腦空間認知和導航具有重要作用,但如何篩選位置細胞尚缺乏系統研究。針對位置細胞篩選,以鴿子為模式動物,利用自由活動狀態下鴿子海馬區鋒電位和運動軌跡信號,給出了鴿子位置細胞的篩選流程,分析了位置細胞篩選過程中網格數和濾波核尺寸對于位置細胞位置野的影響。實測和仿真數據結果表明,該篩選方法可以有效地篩選出位置細胞,位置細胞位置野大小在整體趨勢上與劃分網格數基本呈反比,與高斯濾波核尺寸基本呈正比。該研究不僅能夠促進鴿子位置細胞響應特性研究,而且有助于空間認知與導航神經機制的解析。
引用本文: 劉新玉, 崔書華, 楊晨光, 王東云, 劉凱歌, 秦月, 溫盛軍. 鴿子位置細胞篩選及影響因素分析. 生物醫學工程學雜志, 2024, 41(2): 335-341. doi: 10.7507/1001-5515.202307023 復制
版權信息: ?四川大學華西醫院華西期刊社《生物醫學工程學雜志》版權所有,未經授權不得轉載、改編
0 引言
空間認知與導航是大腦的基本功能,對于個體生存及日常活動具有重要意義。自從美國心理學家Tolman[1]受老鼠迷宮探索實驗啟發首次提出“空間認知地圖”并得到O’Keefe等[2]實驗證實以來,越來越多的結果表明位置細胞在大腦空間認知和導航中起到了關鍵作用。當動物到達空間中特定位置時,大腦海馬區錐體神經元會出現特異性放電,這種特異性放電編碼了動物的位置信息,O’Keefe等[2]將這種具有特異性放電的細胞定義為位置細胞,特異性放電對應的區域定義為位置細胞位置野。位置細胞篩選對于空間認知與導航機制解析具有重要意義,也有助于推動仿生導航相關技術的發展。
隨著研究的深入,位置細胞已在不同動物大腦中被發現。1971年,O’Keefe等[3]首次在大鼠海馬區發現了位置細胞,并對位置細胞位置野特性進行了深入研究,開啟了空間認知與導航機制解析的新紀元;2007年,Ulanovsky等[4]在二維空間中篩選了蝙蝠的位置細胞,發現二維空間中蝙蝠和大鼠的位置細胞位置野特性幾乎沒有任何區別;2013年,Yartsev等[5]進一步對三維空間蝙蝠飛行態時位置細胞及其位置野特性進行了分析;2017年,劉新玉等[6]分析了鴿子海馬區位置細胞位置野的空間響應特性,發現鴿子海馬區位置細胞一般具有多個位置野,它們分布位置相對集中,覆蓋區域大部分都有重疊;2021年,Payne等[7]研究了山雀海馬區位置細胞特性,發現具有食物儲藏能力的山雀位置細胞位置野特性與哺乳類更為相似,而且海馬區前半部分位置細胞位置野更明顯。但是,目前研究人員更多地關注了后端位置細胞響應特性和編碼機制,關于前端位置細胞的篩選研究相對較少。
篩選出神經元的位置野是確定位置細胞的標志。位置野篩選需要經過采樣頻率同步、網格劃分、信號量化、二維濾波、閾值設定及響應區域確定等一系列的步驟,其中每一步參數設置的不同都會造成位置野尺寸和位置的巨大差異,也會直接影響后續位置細胞響應特性的分析。本研究以鴿子為模式動物,在分析其海馬區神經元信號發放特性的基礎上,給出了位置細胞位置野確定的流程和偽代碼,并研究了信號量化中網格數的劃分和信號濾波中濾波核的尺寸等參數對位置細胞篩選的影響,以期提高位置細胞篩選的客觀性和可靠性,并為后續位置細胞位置野特性的研究奠定基礎。
1 材料與方法
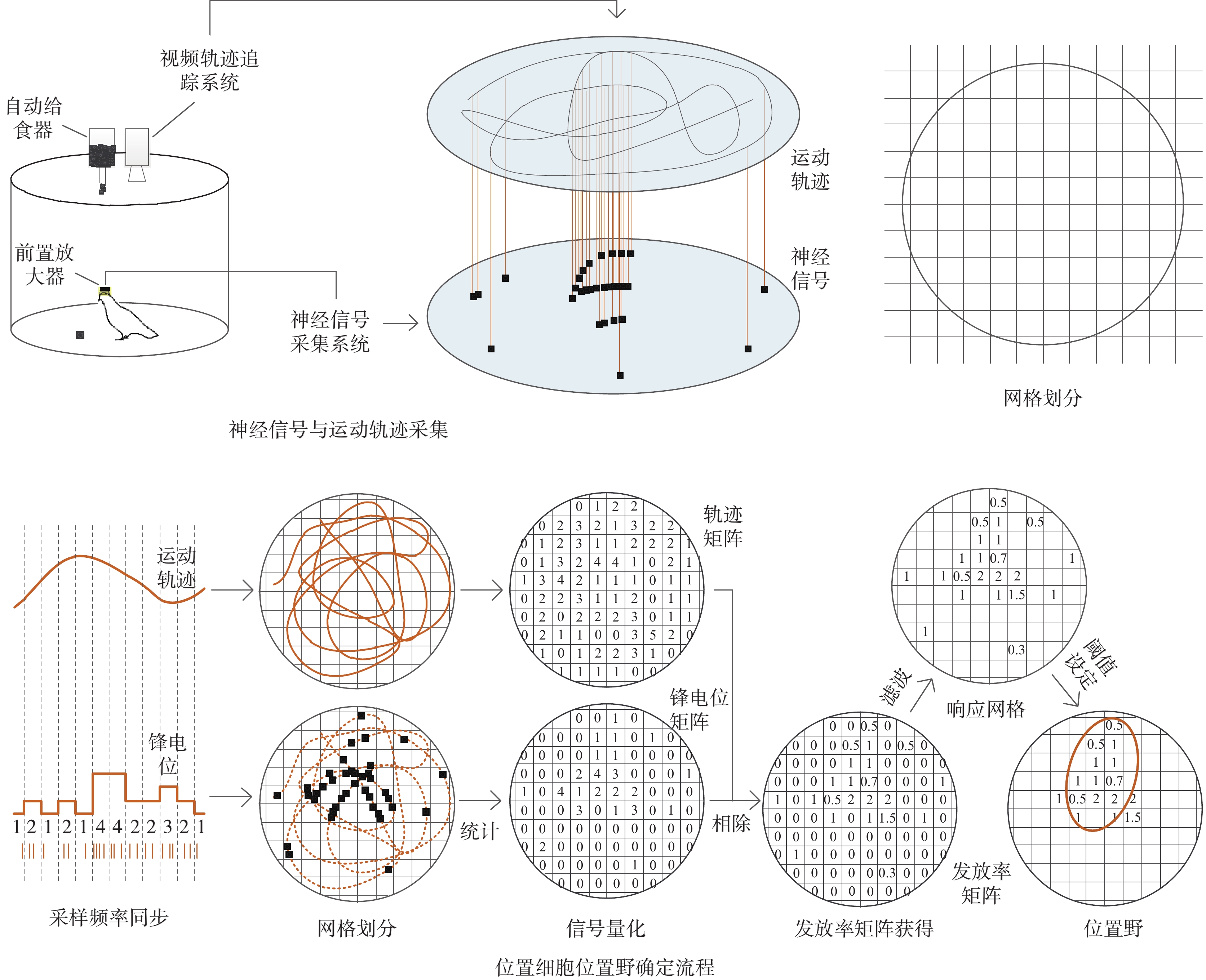
位置細胞的篩選需要大腦神經元鋒電位(spike)信號和與該信號同步采集的動物運動軌跡。在篩選過程中首先將動物的活動區域劃分為不同的網格數,然后計算每個網格中動物經過的次數和神經元spike發放的個數,將兩者相除即可得到spike發放率分布圖,根據spike發放率的大小確定出響應網格即位置野,以篩選出位置細胞。
1.1 信號采集與處理
信號采集以預訓練篩選后的成年鴿子(n = 4,體重450~550 g,雌雄各半)為對象,信號采集前需要在保證鴿子自由飲水情況下控制飲食,以增強動物在信號采集中覓食的動力。記錄鴿子的體重變化,待體重變為正常體重的80%左右時在直徑1 m的圓桶中進行自由覓食(如圖1所示)。實驗圓桶上方有一自動給食裝置,每間隔10 s下落一粒食物(黃豆),引誘鴿子在圓桶中自主運動,以使運動軌跡遍布整個實驗場。每天進行一次信號采集,包含6個試次,每個試次持續5 min。每只鴿子記錄一周左右。
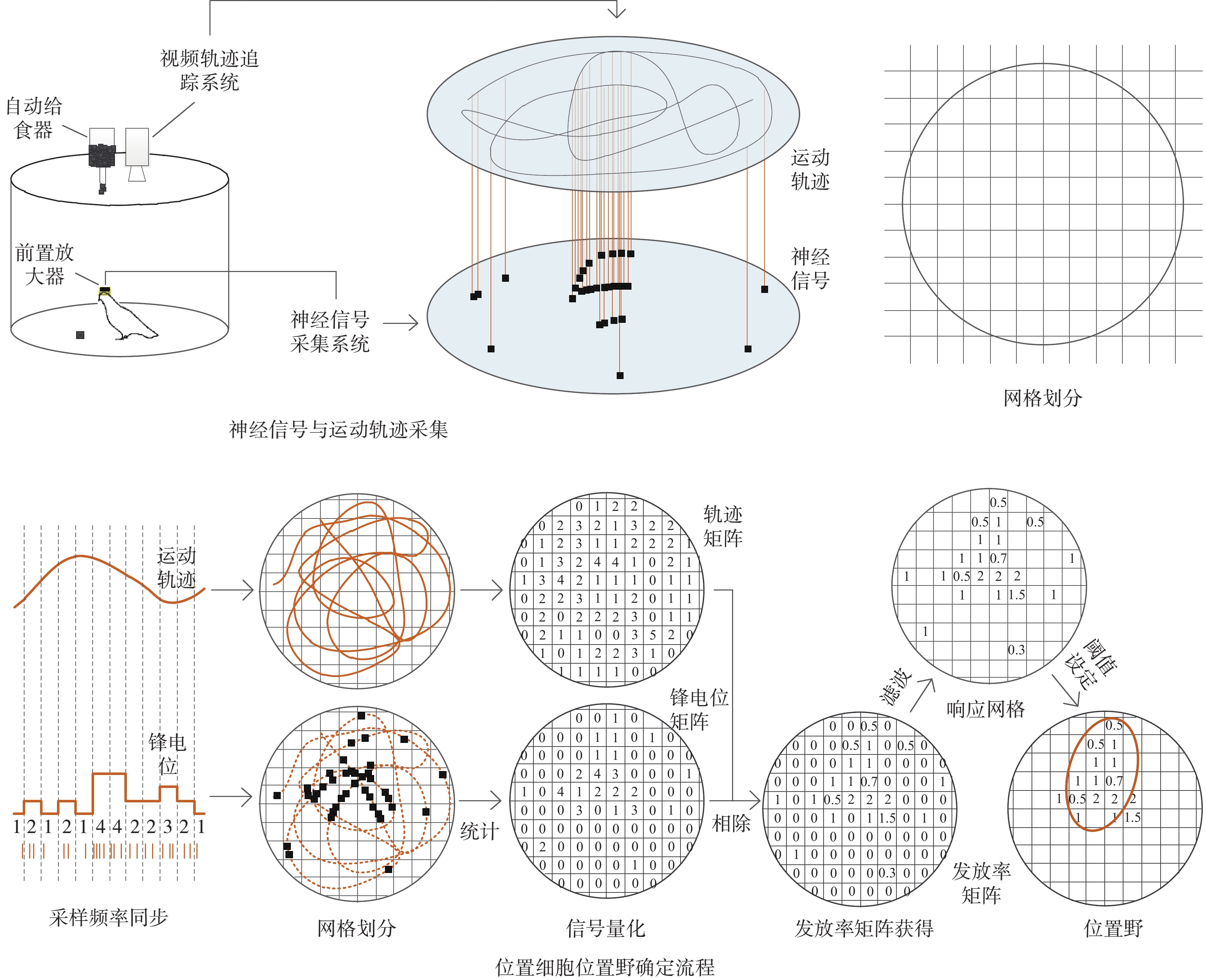 圖1
信號采集及位置細胞篩選流程示意圖
Figure1.
Schematic diagram of signal acquisition and place cell screening process
圖1
信號采集及位置細胞篩選流程示意圖
Figure1.
Schematic diagram of signal acquisition and place cell screening process
信號采集采用標準的胞外微電極陣列植入技術[8],術前根據動物的體重用3%戊巴比妥那(0.12 mL/100 g)進行全身麻醉,在頭部皮膚開口處用2%利多卡因進行局部麻醉以減少動物疼痛,麻醉后將鴿子俯臥固定在定制的腦立體定位儀(68505輕便型,深圳市瑞沃德生命科技有限公司,中國)上[9]。腦區定位參考鴿子腦功能圖譜[10]確定Hp區位置(AP:4.0~7.5 mm,ML: 0~3 mm,DP: 0~3 mm),去除顱骨和硬腦膜后進行微電極(微針陣列電極,16通道,4×4排列,鎢絲,直徑100 μm,陣列間距200 μm,蘇州科斗腦機科技有限公司,中國)植入,植入完成后用牙科水泥進行固定,待牙科水泥固化后進行傷口縫合和消毒處理,等待鴿子蘇醒和康復。
待鴿子5~7天恢復后,在圓桶內鴿子自由覓食時使用多通道神經信號記錄系統(ZeusTM,Bio-Signal Technologies,美國)進行信號采集(采樣頻率30 kHz),同時記錄鴿子的運動軌跡(CyclopsTM,江蘇易格生物科技有限公司,中國)。實驗分別記錄了3只鴿子的神經信號和軌跡數據,分別編號為026、029、060。采集的神經信號經過二階巴特沃斯(Butterworth)濾波器濾波(0.25~5 kHz)、幅度閾值法檢測、電極間相關法去噪等步驟,獲得不同通道的spike發放序列。
此外,為了測試位置細胞篩選方法的可靠性,文中也使用了部分仿真數據。仿真數據采用已發表的文獻數據,數據包含時長24.3 min的49通道數據,采樣頻率為30 kHz,具體參數請參考文獻[11]。
1.2 位置細胞位置野確定
確定位置細胞位置野的過程也就是篩選位置細胞的過程。這個過程中需要經過采樣頻率同步、網格劃分、信號量化、發放率矩陣獲得、二維濾波、閾值設定及響應區域確定等步驟,如圖1所示。為了能更好地描述位置細胞位置野的確定過程,假設在T時間段內共采集到神經元spike個數為N個、動物位置為M個,則神經元spike序列可以表示為N = {N(t), t = 0:1/fspike:T},其中fspike為spike信號的采樣頻率,動物運動軌跡可以表示為M = {(x(t),y(t)), t = 0:1/fpath:T},其中fpath為運動軌跡的采樣頻率,x(t)為運動軌跡x軸的位置,y(t)為運動軌跡y軸的位置。由于神經信號和運動軌跡的采樣頻率不同,因此在數據量化之前首先需要將spike序列與運動軌跡對齊,即統計每個運動位置上發放的spike數量。具體如下:
 '/> '/> |
其中ti = i × 1/fpath,i = 0:T × fpath,則  = {
= { , t = 0:1/fpath:T }。
, t = 0:1/fpath:T }。
信號對齊后就是信號的量化,即將實驗場等值劃分為一定的網格,計算網格內動物運動位置和spike信號的個數。假設將二維平面劃分為n×m個網格,計算每個網格中運動位置和spike個數,獲得spike數據矩陣Rs和軌跡發放矩陣Rp。
信號量化完成后,將spike數據矩陣和軌跡數據矩陣相除就可以獲得神經元在不同網格中的spike發放率,即spike發放率矩陣Rf,計算如下:
 |
上式中  代表兩矩陣中的每個元素相除,如果Rs(i,j) = 0且Rp(i,j) = 0則Rf(i,j) = 0。
代表兩矩陣中的每個元素相除,如果Rs(i,j) = 0且Rp(i,j) = 0則Rf(i,j) = 0。
發放率矩陣Rf一般會存在很強噪聲干擾,導致有些網格的值異常大,降低了位置野確定的難度。為了去除發放率矩陣中的奇異值,采用文獻中常用的二維高斯濾波算法對發放率矩陣進行濾波處理,具體計算公式如下[6]:
 |
其中IG(i, j)為濾波后的值,Rf(i + p, j + q)是發放率矩陣數值,k為濾波器尺寸,σ為高斯函數的標準差。
為了量化位置野尺寸,利用3σ準則,即計算發放率矩陣Rf的均值μ與方差σ,篩選出spike發放率滿足Rf(i, j) > μ + σ時的網格,定義這些網格為響應網格。若滿足連通的響應網格數  ,則定義此響應網格的區域為位置細胞位置野[6]。假設已知位置細胞spike發放時間數據spktime、包含x軸和y軸坐標點的軌跡數據pathdata以及軌跡數據的采樣時間數據pathtime,位置細胞篩選核心步驟的MATLAB偽代碼如下:
,則定義此響應網格的區域為位置細胞位置野[6]。假設已知位置細胞spike發放時間數據spktime、包含x軸和y軸坐標點的軌跡數據pathdata以及軌跡數據的采樣時間數據pathtime,位置細胞篩選核心步驟的MATLAB偽代碼如下:
1.3 位置細胞篩選影響因素
由位置野的確定過程可知,位置細胞的篩選受兩個因素的影響,即網格劃分數和二維高斯濾波的濾波核尺寸。因為在位置細胞篩選過程中只有這兩個步驟中的參數是不固定的,盡管不同文獻中設定了不同的參數[12-13],但很少有文獻討論這些參數是如何影響位置細胞位置野的。
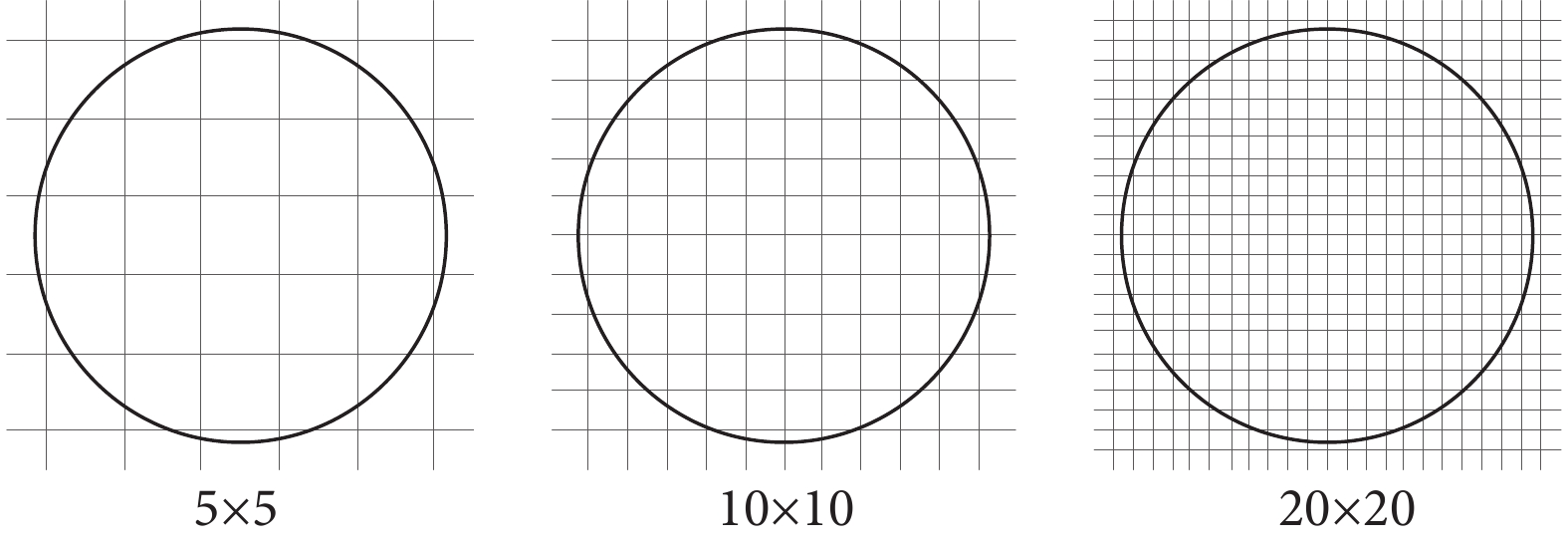
(1)網格劃分數的影響:網格劃分是信號量化中的核心環節。為了計算不同位置spike信號的發放率,以確定位置野的位置和尺寸,就需要對實驗場劃分不同的網格,如圖2所示。為了量化網格數劃分對位置野的影響,我們計算了劃分不同網格數時得到的響應網格面積。
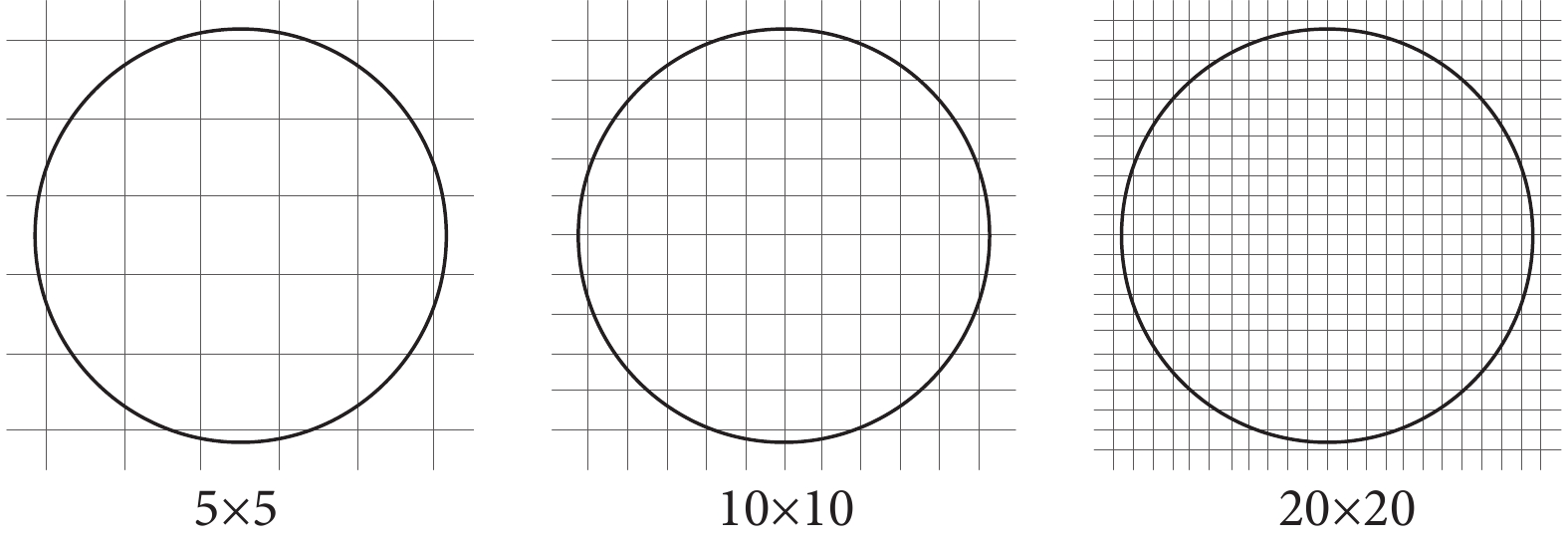 圖2
不同網格數劃分示意
Figure2.
Schematic diagram for dividing different grid numbers
圖2
不同網格數劃分示意
Figure2.
Schematic diagram for dividing different grid numbers
(2)濾波核尺寸的影響:為了減少奇異值對位置野確定的影響以及準確地確定位置野的尺寸和位置,濾波是必不可少的步驟。在常用的二維高斯濾波中,濾波核尺寸是核心參數,影響著濾波后位置細胞篩選的效果。為了探討濾波核尺寸對位置細胞位置野的影響,我們計算了隨著濾波核尺寸改變位置細胞響應網格面積的變化趨勢。
2 結果
為了篩選鴿子海馬區位置細胞,并探究位置細胞篩選過程中的影響因素,利用仿真數據和在鴿子海馬區獲得的實測數據從篩選的位置細胞位置野、劃分網格數對位置野的影響、濾波核尺寸對位置野的影響等三個方面進行了論述。
2.1 篩選的位置細胞位置野
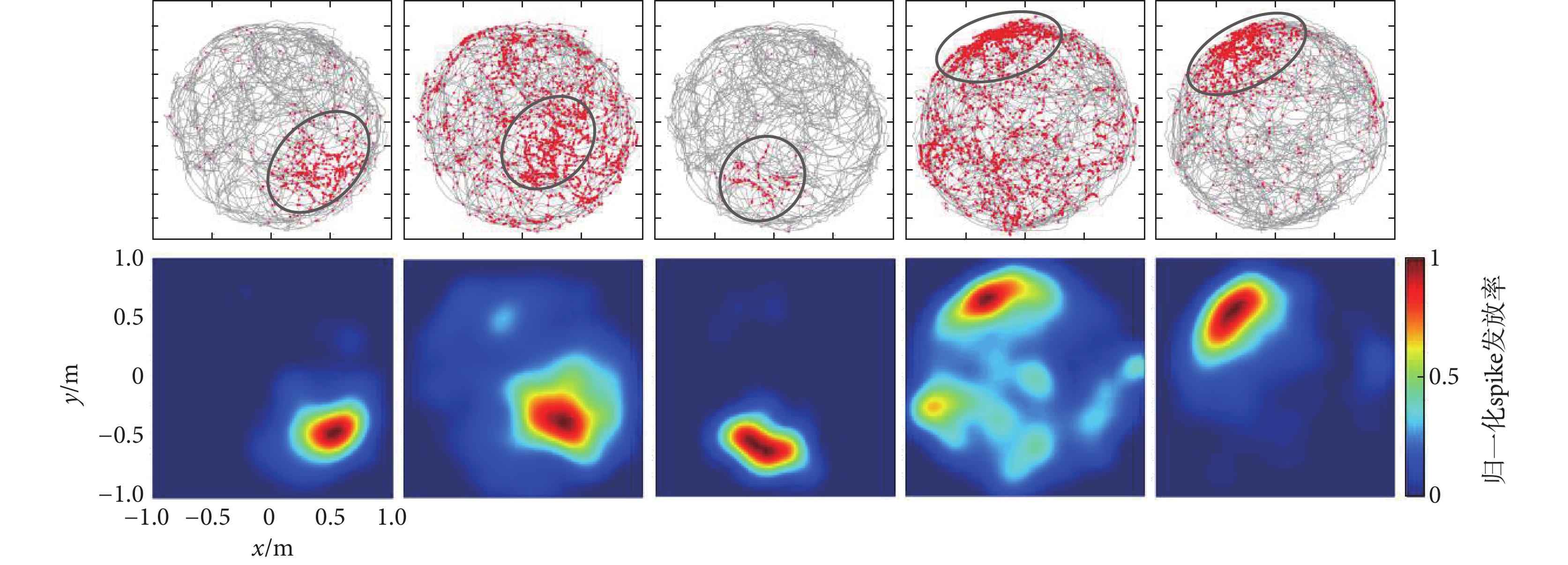
仿真數據選擇已發表的實驗數據,包含時長24.3 min的49通道數據,圖3給出了篩選出來的5個典型位置細胞位置野。圖3上圖為位置細胞spike發放分布圖,圖中每個紅色的圓點代表一個spike,灰色的線條為動物運動軌跡,spike即紅色圓點聚集的區域即為位置細胞位置野;圖3下圖為確定的位置細胞位置野,圖中紅色越深代表spike發放率越大,藍色越深代表spike發放率越小,圖中每個位置細胞的spike發放率都進行了歸一化處理。
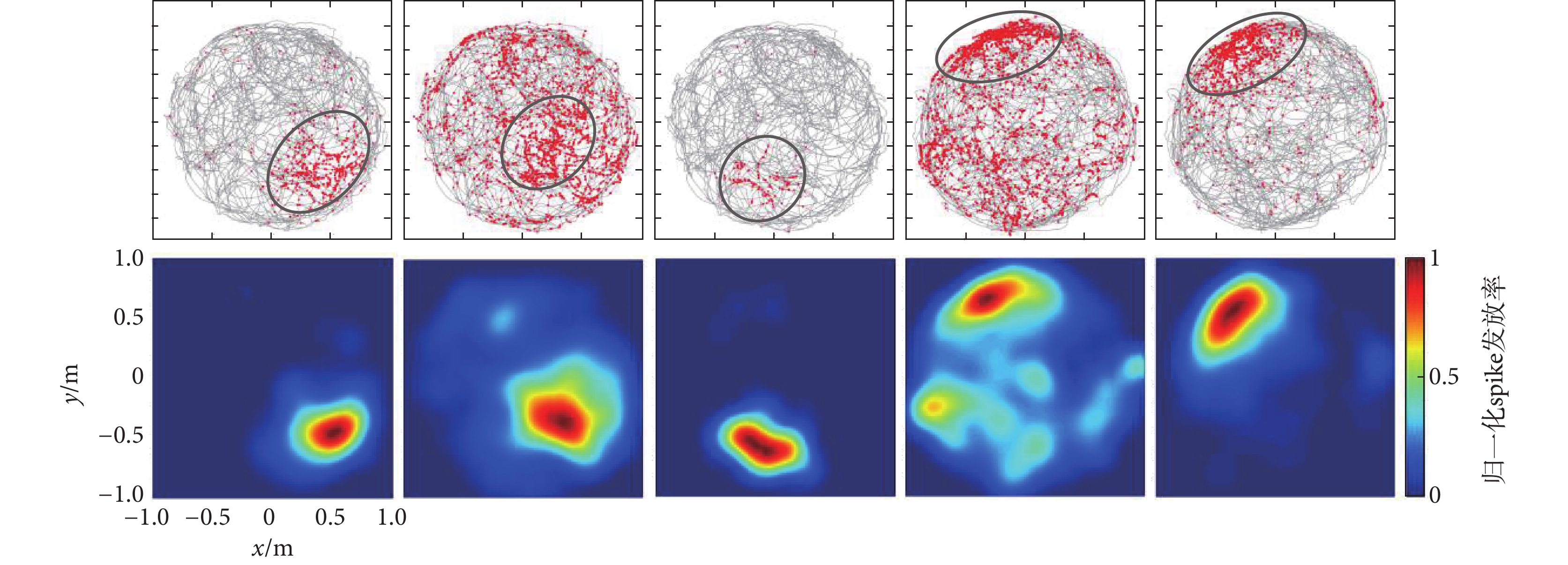 圖3
仿真數據篩選的位置細胞及其位置野
Figure3.
Place cells and its place fields screened from simulation data
圖3
仿真數據篩選的位置細胞及其位置野
Figure3.
Place cells and its place fields screened from simulation data
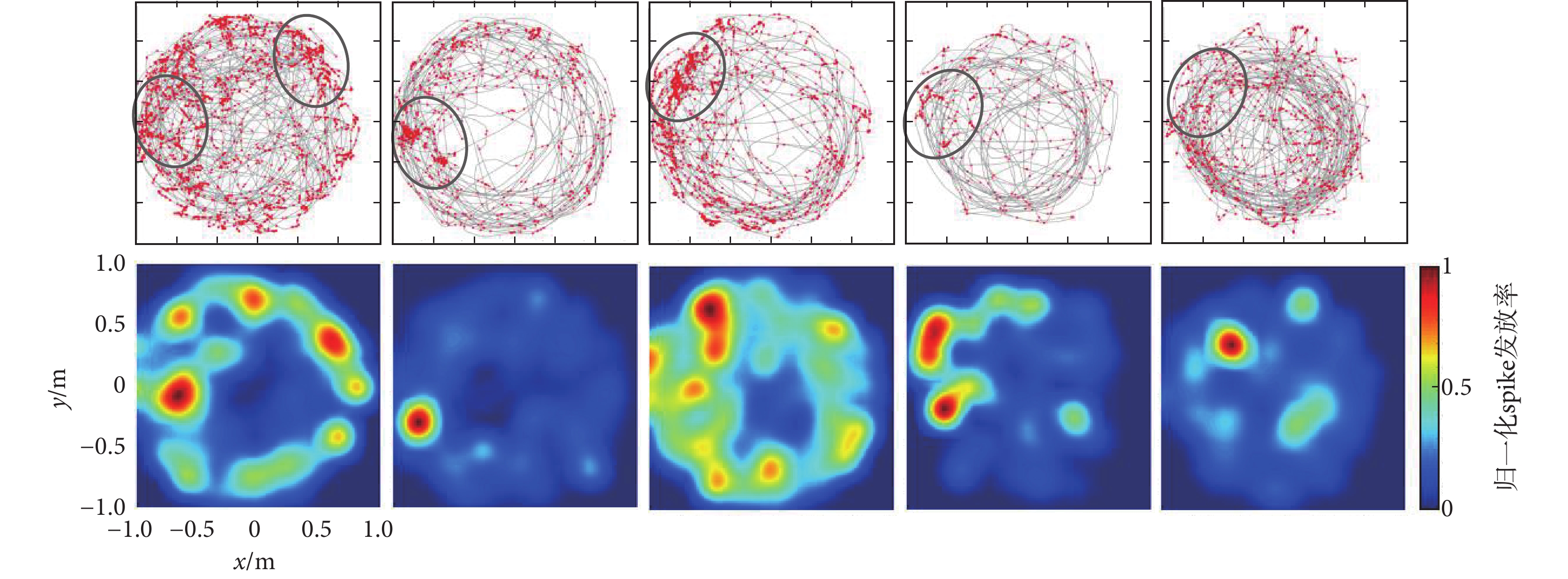
實測數據由鴿子海馬區神經元spike信號和鴿子運動軌跡獲得的位置細胞位置野,圖4為篩選出來的5個典型位置細胞位置野,圖中數據代表的含義與圖3一致。由圖可知,實測的鴿子位置細胞位置野盡管沒有仿真數據編碼區域清晰,但是經過固定的篩選流程,每個位置細胞也顯現出了特定的位置野區域,這一結果表明鴿子海馬區還存在類似于哺乳動物的位置細胞。
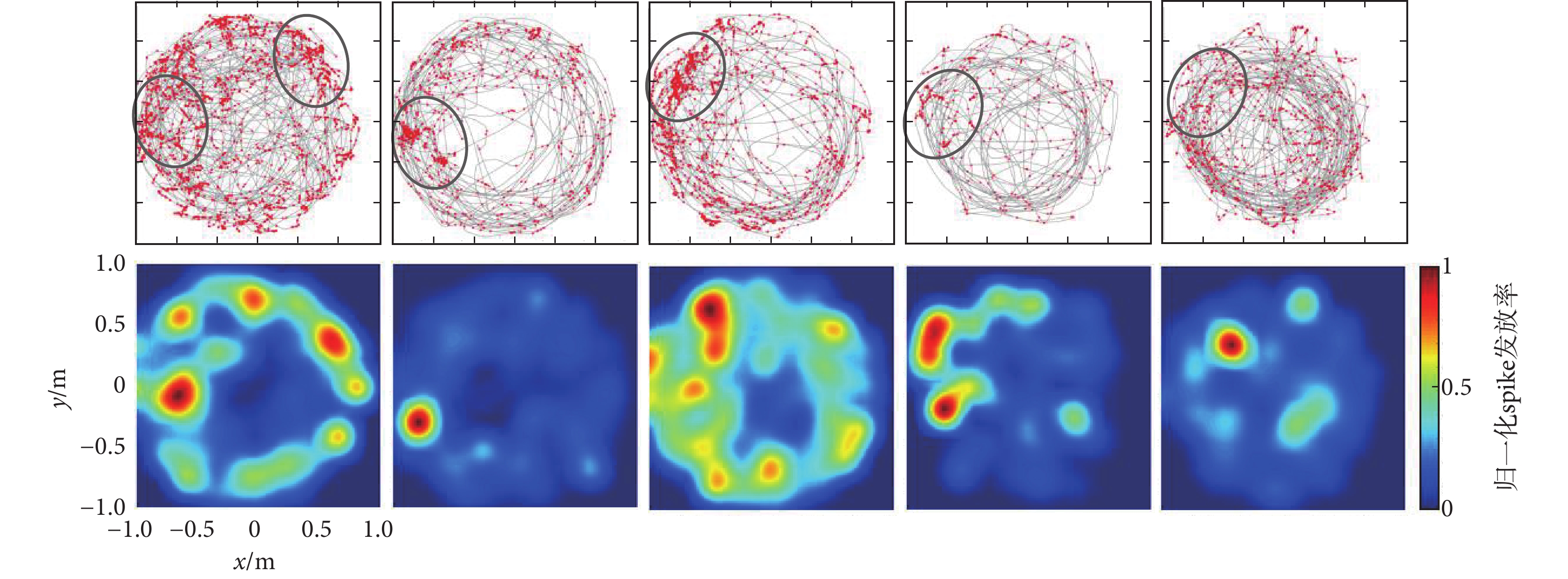 圖4
實測數據篩選的位置細胞及其位置野
Figure4.
Place cells and its place fields screened from real data
圖4
實測數據篩選的位置細胞及其位置野
Figure4.
Place cells and its place fields screened from real data
2.2 劃分網格數對位置野的影響
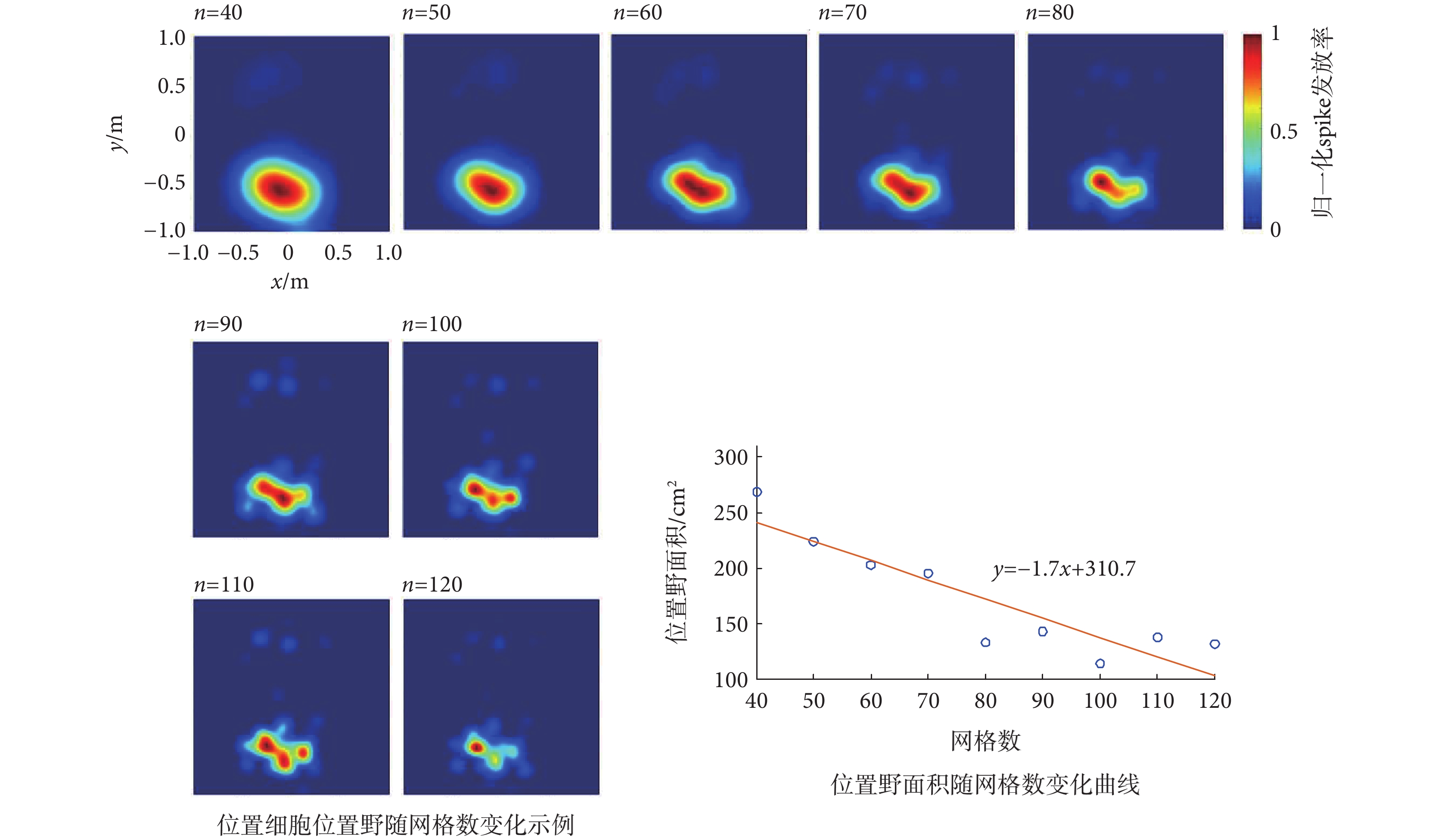
網格數對位置細胞位置野的影響如圖5所示,包括典型的位置細胞位置野隨網格數(40~100)變化的示例,以及位置野面積隨網格數變化的曲線。由圖可知,隨著劃分網格數的逐漸增大,位置野會逐漸縮小,呈明顯的下降趨勢,而且由變化曲線可知,在n≤70時位置野面積下降速度較大,當n > 70時位置野面積則趨于平穩。這一結果表明,網格數的大小會影響位置野的尺寸,劃分網格數應當小于70為宜。
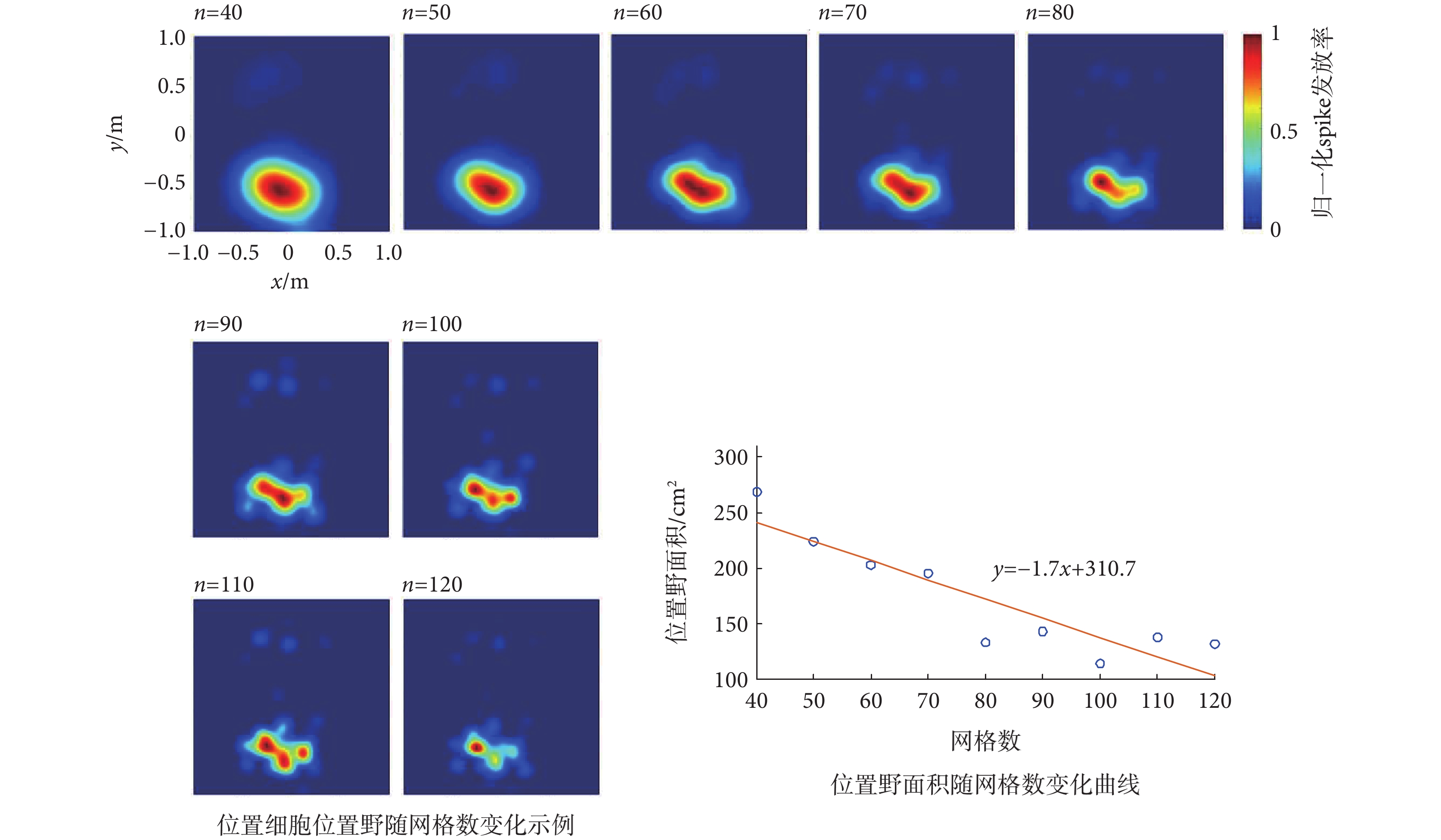 圖5
網格數對位置細胞位置野的影響
Figure5.
Effect of the grid number on the place field of place cell
圖5
網格數對位置細胞位置野的影響
Figure5.
Effect of the grid number on the place field of place cell
2.3 濾波核尺寸對位置野的影響
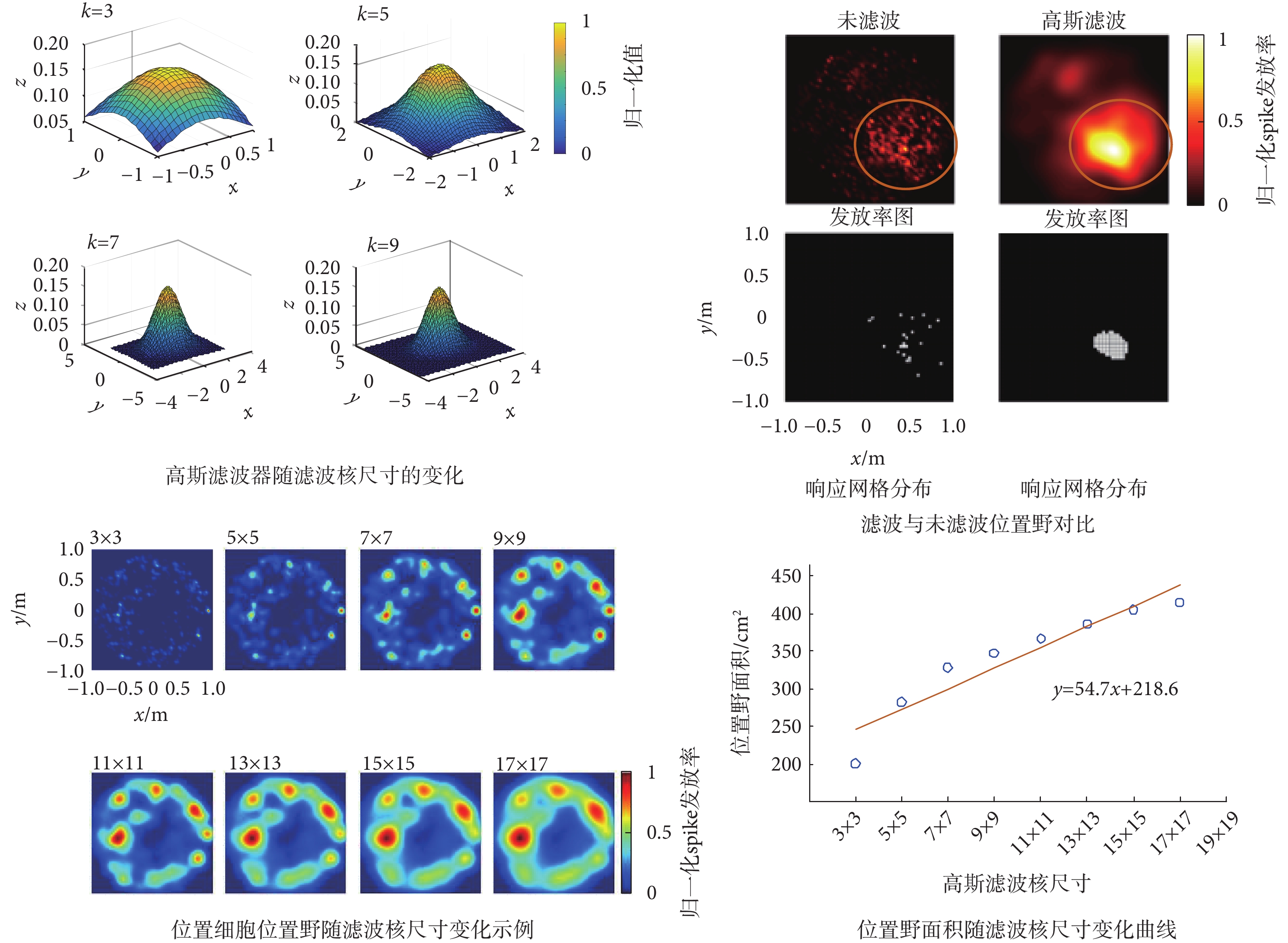
濾波核尺寸對位置細胞位置野的影響如圖6所示。由圖可知,隨著濾波核尺寸的增大濾波器會逐漸變小,而且濾波后位置野會更加明顯。通過對位置野隨濾波核尺寸的變化分析表明,隨著濾波核尺寸的增加,位置細胞位置野面積會逐步增大,當k≤7時位置野增大的趨勢比較明顯,當k > 7時位置野增大的趨勢則趨于平緩。這一結果表明二維高斯濾波器濾波核的尺寸會影響位置野的大小,濾波核尺寸應大于7為宜。
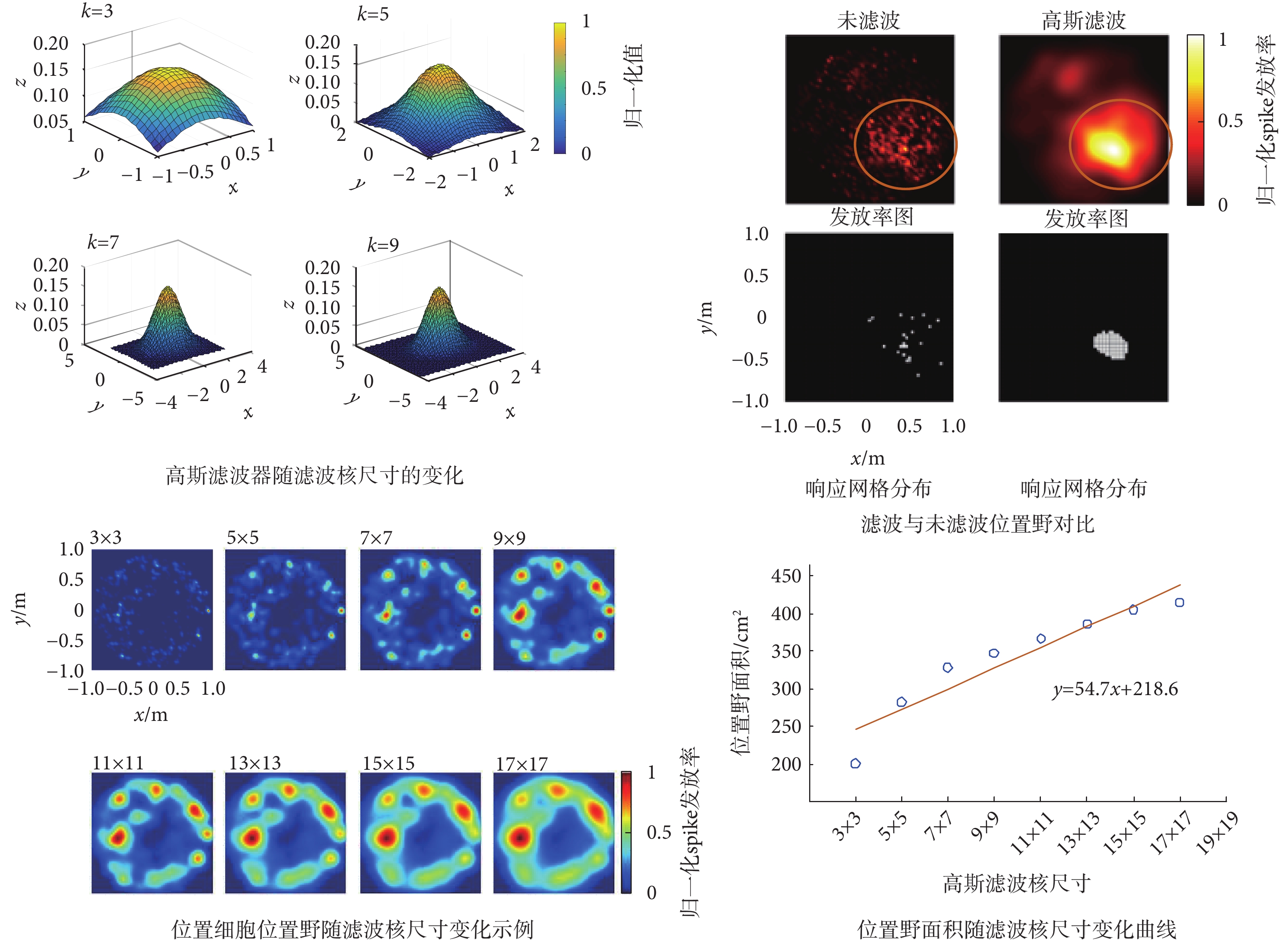 圖6
濾波核尺寸對位置細胞位置野的影響
Figure6.
Effect of the filtering kernel size on the place field of place cell
圖6
濾波核尺寸對位置細胞位置野的影響
Figure6.
Effect of the filtering kernel size on the place field of place cell
3 討論與結論
位置細胞的篩選對于大腦空間認知和導航機制的研究至關重要,但目前未見針對位置細胞篩選的系統研究。本文針對鴿子位置細胞的篩選及其影響因素,給出了位置細胞篩選的具體過程,并分析了網格數和濾波核尺寸對位置細胞位置野的影響。實測與仿真數據結果表明:① 本文給出的位置細胞篩選方法可以有效地確定位置細胞位置野,篩選出位置細胞;② 位置野大小會受到網格數的影響,網格數與位置野面積呈反比;③ 二維高斯濾波中濾波核尺寸會影響位置野的大小,濾波核尺寸與位置野面積呈正比。這些研究結果對于鳥類和其他動物位置細胞的篩選可能會起到積極的促進作用。
位置細胞的篩選與位置野的確定是同一過程,因為只有具有位置野的神經元才會被認為是位置細胞。位置細胞篩選需要動物運動軌跡數據和相應時段的神經元spike信號,然后經過采樣頻率同步、網格劃分、信號量化、發放率矩陣獲得、二維濾波、閾值設定及響應區域確定等步驟以確定是否存在位置野,只有存在位置野的神經元才認為是位置細胞。在位置細胞位置野確定過程中,網格劃分數和濾波核尺寸設定是難點,受參數因素影響較大,需要根據實際情況進行設定,不同的參數設定可能會得出不同的結果。在整體趨勢上,位置野大小與劃分網格數基本呈反比,與高斯濾波核尺寸基本呈正比。就鴿子位置細胞篩選而言,我們認為參數設定以劃分網格數小于70、濾波核尺寸大于7為宜。
此外,位置細胞篩選軌跡數據和spike信號的記錄開始和結束時間及采樣率必須一致。一般情況下,我們很容易實現軌跡數據和spike信號的記錄時間同步,但是它們的采樣率往往是不同的,軌跡數據采樣率(30 Hz)一般會顯著低于spike信號采樣率(30 000 Hz),因此位置細胞篩選之前需要將這兩個信號的采樣率調整一致。需要注意的是,采樣率調整不能通過傳統的降采樣或升采樣的方式進行,而是通過統計每一個軌跡點所處時間區間內spike的個數,使兩種信號的采樣率調整一致。而且在信號采集中,需要使動物軌跡盡可能均勻地布滿整個活動空間,動物運動軌跡布滿活動空間是位置細胞篩選的前提,而運動軌跡均勻地布滿活動空間則是獲得可靠位置野的保障。
位置細胞篩選是探索空間認知與導航神經機制的重要步驟。針對位置細胞的篩選,以鴿子為模式動物,詳細給出了位置細胞的篩選過程,并分析了位置細胞篩選過程中的影響因素。本文的研究結果不僅對鴿子位置細胞篩選具有較高應用價值,而且對其他動物位置細胞的篩選也具有指導意義。理論上,二維平面中不同種類動物大腦位置細胞篩選的方法和流程是一致的,僅僅存在運動方式和活動空間大小的差異。另外,位置細胞的篩選只是解析大腦空間認知與導航機制的基礎和前提,后續還需要進一步解析鳥類是如何在空間中進行認知和導航的,以及鳥類與哺乳類是否分享了相同的空間信息處理機制。
重要聲明
利益沖突聲明:本文全體作者均聲明不存在利益沖突。
作者貢獻聲明:劉新玉負責研究計劃安排、動物實驗手術以及論文的書寫。崔書華負責論文修改稿書寫和實驗數據的處理。楊晨光負責信號的采集和處理。王東云負責研究計劃指導。劉凱歌負責實驗動物飼養和培育。秦月負責實驗數據記錄。溫盛軍負責研究計劃監督和論文修訂。
倫理聲明:本研究通過了黃淮學院倫理委員會的審批(批文編號:LL20220001)。
0 引言
空間認知與導航是大腦的基本功能,對于個體生存及日常活動具有重要意義。自從美國心理學家Tolman[1]受老鼠迷宮探索實驗啟發首次提出“空間認知地圖”并得到O’Keefe等[2]實驗證實以來,越來越多的結果表明位置細胞在大腦空間認知和導航中起到了關鍵作用。當動物到達空間中特定位置時,大腦海馬區錐體神經元會出現特異性放電,這種特異性放電編碼了動物的位置信息,O’Keefe等[2]將這種具有特異性放電的細胞定義為位置細胞,特異性放電對應的區域定義為位置細胞位置野。位置細胞篩選對于空間認知與導航機制解析具有重要意義,也有助于推動仿生導航相關技術的發展。
隨著研究的深入,位置細胞已在不同動物大腦中被發現。1971年,O’Keefe等[3]首次在大鼠海馬區發現了位置細胞,并對位置細胞位置野特性進行了深入研究,開啟了空間認知與導航機制解析的新紀元;2007年,Ulanovsky等[4]在二維空間中篩選了蝙蝠的位置細胞,發現二維空間中蝙蝠和大鼠的位置細胞位置野特性幾乎沒有任何區別;2013年,Yartsev等[5]進一步對三維空間蝙蝠飛行態時位置細胞及其位置野特性進行了分析;2017年,劉新玉等[6]分析了鴿子海馬區位置細胞位置野的空間響應特性,發現鴿子海馬區位置細胞一般具有多個位置野,它們分布位置相對集中,覆蓋區域大部分都有重疊;2021年,Payne等[7]研究了山雀海馬區位置細胞特性,發現具有食物儲藏能力的山雀位置細胞位置野特性與哺乳類更為相似,而且海馬區前半部分位置細胞位置野更明顯。但是,目前研究人員更多地關注了后端位置細胞響應特性和編碼機制,關于前端位置細胞的篩選研究相對較少。
篩選出神經元的位置野是確定位置細胞的標志。位置野篩選需要經過采樣頻率同步、網格劃分、信號量化、二維濾波、閾值設定及響應區域確定等一系列的步驟,其中每一步參數設置的不同都會造成位置野尺寸和位置的巨大差異,也會直接影響后續位置細胞響應特性的分析。本研究以鴿子為模式動物,在分析其海馬區神經元信號發放特性的基礎上,給出了位置細胞位置野確定的流程和偽代碼,并研究了信號量化中網格數的劃分和信號濾波中濾波核的尺寸等參數對位置細胞篩選的影響,以期提高位置細胞篩選的客觀性和可靠性,并為后續位置細胞位置野特性的研究奠定基礎。
1 材料與方法
位置細胞的篩選需要大腦神經元鋒電位(spike)信號和與該信號同步采集的動物運動軌跡。在篩選過程中首先將動物的活動區域劃分為不同的網格數,然后計算每個網格中動物經過的次數和神經元spike發放的個數,將兩者相除即可得到spike發放率分布圖,根據spike發放率的大小確定出響應網格即位置野,以篩選出位置細胞。
1.1 信號采集與處理
信號采集以預訓練篩選后的成年鴿子(n = 4,體重450~550 g,雌雄各半)為對象,信號采集前需要在保證鴿子自由飲水情況下控制飲食,以增強動物在信號采集中覓食的動力。記錄鴿子的體重變化,待體重變為正常體重的80%左右時在直徑1 m的圓桶中進行自由覓食(如圖1所示)。實驗圓桶上方有一自動給食裝置,每間隔10 s下落一粒食物(黃豆),引誘鴿子在圓桶中自主運動,以使運動軌跡遍布整個實驗場。每天進行一次信號采集,包含6個試次,每個試次持續5 min。每只鴿子記錄一周左右。
圖1
信號采集及位置細胞篩選流程示意圖
Figure1.
Schematic diagram of signal acquisition and place cell screening process
信號采集采用標準的胞外微電極陣列植入技術[8],術前根據動物的體重用3%戊巴比妥那(0.12 mL/100 g)進行全身麻醉,在頭部皮膚開口處用2%利多卡因進行局部麻醉以減少動物疼痛,麻醉后將鴿子俯臥固定在定制的腦立體定位儀(68505輕便型,深圳市瑞沃德生命科技有限公司,中國)上[9]。腦區定位參考鴿子腦功能圖譜[10]確定Hp區位置(AP:4.0~7.5 mm,ML: 0~3 mm,DP: 0~3 mm),去除顱骨和硬腦膜后進行微電極(微針陣列電極,16通道,4×4排列,鎢絲,直徑100 μm,陣列間距200 μm,蘇州科斗腦機科技有限公司,中國)植入,植入完成后用牙科水泥進行固定,待牙科水泥固化后進行傷口縫合和消毒處理,等待鴿子蘇醒和康復。
待鴿子5~7天恢復后,在圓桶內鴿子自由覓食時使用多通道神經信號記錄系統(ZeusTM,Bio-Signal Technologies,美國)進行信號采集(采樣頻率30 kHz),同時記錄鴿子的運動軌跡(CyclopsTM,江蘇易格生物科技有限公司,中國)。實驗分別記錄了3只鴿子的神經信號和軌跡數據,分別編號為026、029、060。采集的神經信號經過二階巴特沃斯(Butterworth)濾波器濾波(0.25~5 kHz)、幅度閾值法檢測、電極間相關法去噪等步驟,獲得不同通道的spike發放序列。
此外,為了測試位置細胞篩選方法的可靠性,文中也使用了部分仿真數據。仿真數據采用已發表的文獻數據,數據包含時長24.3 min的49通道數據,采樣頻率為30 kHz,具體參數請參考文獻[11]。
1.2 位置細胞位置野確定
確定位置細胞位置野的過程也就是篩選位置細胞的過程。這個過程中需要經過采樣頻率同步、網格劃分、信號量化、發放率矩陣獲得、二維濾波、閾值設定及響應區域確定等步驟,如圖1所示。為了能更好地描述位置細胞位置野的確定過程,假設在T時間段內共采集到神經元spike個數為N個、動物位置為M個,則神經元spike序列可以表示為N = {N(t), t = 0:1/fspike:T},其中fspike為spike信號的采樣頻率,動物運動軌跡可以表示為M = {(x(t),y(t)), t = 0:1/fpath:T},其中fpath為運動軌跡的采樣頻率,x(t)為運動軌跡x軸的位置,y(t)為運動軌跡y軸的位置。由于神經信號和運動軌跡的采樣頻率不同,因此在數據量化之前首先需要將spike序列與運動軌跡對齊,即統計每個運動位置上發放的spike數量。具體如下:
| '/> |
其中ti = i × 1/fpath,i = 0:T × fpath,則 = {, t = 0:1/fpath:T }。
信號對齊后就是信號的量化,即將實驗場等值劃分為一定的網格,計算網格內動物運動位置和spike信號的個數。假設將二維平面劃分為n×m個網格,計算每個網格中運動位置和spike個數,獲得spike數據矩陣Rs和軌跡發放矩陣Rp。
信號量化完成后,將spike數據矩陣和軌跡數據矩陣相除就可以獲得神經元在不同網格中的spike發放率,即spike發放率矩陣Rf,計算如下:
|
上式中 代表兩矩陣中的每個元素相除,如果Rs(i,j) = 0且Rp(i,j) = 0則Rf(i,j) = 0。
發放率矩陣Rf一般會存在很強噪聲干擾,導致有些網格的值異常大,降低了位置野確定的難度。為了去除發放率矩陣中的奇異值,采用文獻中常用的二維高斯濾波算法對發放率矩陣進行濾波處理,具體計算公式如下[6]:
|
其中IG(i, j)為濾波后的值,Rf(i + p, j + q)是發放率矩陣數值,k為濾波器尺寸,σ為高斯函數的標準差。
為了量化位置野尺寸,利用3σ準則,即計算發放率矩陣Rf的均值μ與方差σ,篩選出spike發放率滿足Rf(i, j) > μ + σ時的網格,定義這些網格為響應網格。若滿足連通的響應網格數 ,則定義此響應網格的區域為位置細胞位置野[6]。假設已知位置細胞spike發放時間數據spktime、包含x軸和y軸坐標點的軌跡數據pathdata以及軌跡數據的采樣時間數據pathtime,位置細胞篩選核心步驟的MATLAB偽代碼如下:
1.3 位置細胞篩選影響因素
由位置野的確定過程可知,位置細胞的篩選受兩個因素的影響,即網格劃分數和二維高斯濾波的濾波核尺寸。因為在位置細胞篩選過程中只有這兩個步驟中的參數是不固定的,盡管不同文獻中設定了不同的參數[12-13],但很少有文獻討論這些參數是如何影響位置細胞位置野的。
(1)網格劃分數的影響:網格劃分是信號量化中的核心環節。為了計算不同位置spike信號的發放率,以確定位置野的位置和尺寸,就需要對實驗場劃分不同的網格,如圖2所示。為了量化網格數劃分對位置野的影響,我們計算了劃分不同網格數時得到的響應網格面積。
圖2
不同網格數劃分示意
Figure2.
Schematic diagram for dividing different grid numbers
(2)濾波核尺寸的影響:為了減少奇異值對位置野確定的影響以及準確地確定位置野的尺寸和位置,濾波是必不可少的步驟。在常用的二維高斯濾波中,濾波核尺寸是核心參數,影響著濾波后位置細胞篩選的效果。為了探討濾波核尺寸對位置細胞位置野的影響,我們計算了隨著濾波核尺寸改變位置細胞響應網格面積的變化趨勢。
2 結果
為了篩選鴿子海馬區位置細胞,并探究位置細胞篩選過程中的影響因素,利用仿真數據和在鴿子海馬區獲得的實測數據從篩選的位置細胞位置野、劃分網格數對位置野的影響、濾波核尺寸對位置野的影響等三個方面進行了論述。
2.1 篩選的位置細胞位置野
仿真數據選擇已發表的實驗數據,包含時長24.3 min的49通道數據,圖3給出了篩選出來的5個典型位置細胞位置野。圖3上圖為位置細胞spike發放分布圖,圖中每個紅色的圓點代表一個spike,灰色的線條為動物運動軌跡,spike即紅色圓點聚集的區域即為位置細胞位置野;圖3下圖為確定的位置細胞位置野,圖中紅色越深代表spike發放率越大,藍色越深代表spike發放率越小,圖中每個位置細胞的spike發放率都進行了歸一化處理。
圖3
仿真數據篩選的位置細胞及其位置野
Figure3.
Place cells and its place fields screened from simulation data
實測數據由鴿子海馬區神經元spike信號和鴿子運動軌跡獲得的位置細胞位置野,圖4為篩選出來的5個典型位置細胞位置野,圖中數據代表的含義與圖3一致。由圖可知,實測的鴿子位置細胞位置野盡管沒有仿真數據編碼區域清晰,但是經過固定的篩選流程,每個位置細胞也顯現出了特定的位置野區域,這一結果表明鴿子海馬區還存在類似于哺乳動物的位置細胞。
圖4
實測數據篩選的位置細胞及其位置野
Figure4.
Place cells and its place fields screened from real data
2.2 劃分網格數對位置野的影響
網格數對位置細胞位置野的影響如圖5所示,包括典型的位置細胞位置野隨網格數(40~100)變化的示例,以及位置野面積隨網格數變化的曲線。由圖可知,隨著劃分網格數的逐漸增大,位置野會逐漸縮小,呈明顯的下降趨勢,而且由變化曲線可知,在n≤70時位置野面積下降速度較大,當n > 70時位置野面積則趨于平穩。這一結果表明,網格數的大小會影響位置野的尺寸,劃分網格數應當小于70為宜。
圖5
網格數對位置細胞位置野的影響
Figure5.
Effect of the grid number on the place field of place cell
2.3 濾波核尺寸對位置野的影響
濾波核尺寸對位置細胞位置野的影響如圖6所示。由圖可知,隨著濾波核尺寸的增大濾波器會逐漸變小,而且濾波后位置野會更加明顯。通過對位置野隨濾波核尺寸的變化分析表明,隨著濾波核尺寸的增加,位置細胞位置野面積會逐步增大,當k≤7時位置野增大的趨勢比較明顯,當k > 7時位置野增大的趨勢則趨于平緩。這一結果表明二維高斯濾波器濾波核的尺寸會影響位置野的大小,濾波核尺寸應大于7為宜。
圖6
濾波核尺寸對位置細胞位置野的影響
Figure6.
Effect of the filtering kernel size on the place field of place cell
3 討論與結論
位置細胞的篩選對于大腦空間認知和導航機制的研究至關重要,但目前未見針對位置細胞篩選的系統研究。本文針對鴿子位置細胞的篩選及其影響因素,給出了位置細胞篩選的具體過程,并分析了網格數和濾波核尺寸對位置細胞位置野的影響。實測與仿真數據結果表明:① 本文給出的位置細胞篩選方法可以有效地確定位置細胞位置野,篩選出位置細胞;② 位置野大小會受到網格數的影響,網格數與位置野面積呈反比;③ 二維高斯濾波中濾波核尺寸會影響位置野的大小,濾波核尺寸與位置野面積呈正比。這些研究結果對于鳥類和其他動物位置細胞的篩選可能會起到積極的促進作用。
位置細胞的篩選與位置野的確定是同一過程,因為只有具有位置野的神經元才會被認為是位置細胞。位置細胞篩選需要動物運動軌跡數據和相應時段的神經元spike信號,然后經過采樣頻率同步、網格劃分、信號量化、發放率矩陣獲得、二維濾波、閾值設定及響應區域確定等步驟以確定是否存在位置野,只有存在位置野的神經元才認為是位置細胞。在位置細胞位置野確定過程中,網格劃分數和濾波核尺寸設定是難點,受參數因素影響較大,需要根據實際情況進行設定,不同的參數設定可能會得出不同的結果。在整體趨勢上,位置野大小與劃分網格數基本呈反比,與高斯濾波核尺寸基本呈正比。就鴿子位置細胞篩選而言,我們認為參數設定以劃分網格數小于70、濾波核尺寸大于7為宜。
此外,位置細胞篩選軌跡數據和spike信號的記錄開始和結束時間及采樣率必須一致。一般情況下,我們很容易實現軌跡數據和spike信號的記錄時間同步,但是它們的采樣率往往是不同的,軌跡數據采樣率(30 Hz)一般會顯著低于spike信號采樣率(30 000 Hz),因此位置細胞篩選之前需要將這兩個信號的采樣率調整一致。需要注意的是,采樣率調整不能通過傳統的降采樣或升采樣的方式進行,而是通過統計每一個軌跡點所處時間區間內spike的個數,使兩種信號的采樣率調整一致。而且在信號采集中,需要使動物軌跡盡可能均勻地布滿整個活動空間,動物運動軌跡布滿活動空間是位置細胞篩選的前提,而運動軌跡均勻地布滿活動空間則是獲得可靠位置野的保障。
位置細胞篩選是探索空間認知與導航神經機制的重要步驟。針對位置細胞的篩選,以鴿子為模式動物,詳細給出了位置細胞的篩選過程,并分析了位置細胞篩選過程中的影響因素。本文的研究結果不僅對鴿子位置細胞篩選具有較高應用價值,而且對其他動物位置細胞的篩選也具有指導意義。理論上,二維平面中不同種類動物大腦位置細胞篩選的方法和流程是一致的,僅僅存在運動方式和活動空間大小的差異。另外,位置細胞的篩選只是解析大腦空間認知與導航機制的基礎和前提,后續還需要進一步解析鳥類是如何在空間中進行認知和導航的,以及鳥類與哺乳類是否分享了相同的空間信息處理機制。
重要聲明
利益沖突聲明:本文全體作者均聲明不存在利益沖突。
作者貢獻聲明:劉新玉負責研究計劃安排、動物實驗手術以及論文的書寫。崔書華負責論文修改稿書寫和實驗數據的處理。楊晨光負責信號的采集和處理。王東云負責研究計劃指導。劉凱歌負責實驗動物飼養和培育。秦月負責實驗數據記錄。溫盛軍負責研究計劃監督和論文修訂。
倫理聲明:本研究通過了黃淮學院倫理委員會的審批(批文編號:LL20220001)。