引用本文: 劉卓, 趙崢, 張亮亮, 楊翠英, 靳兆. 肺纖維化與鐵死亡的相關性分析與機制探討. 華西醫學, 2024, 39(8): 1238-1245. doi: 10.7507/1002-0179.202310269 復制
版權信息: ?四川大學華西醫院華西期刊社《華西醫學》版權所有,未經授權不得轉載、改編
特發性肺纖維化(idiopathic pulmonary fibrosis, IPF)是一種進行性、慢性、不可逆的肺部疾病,其癥狀包括肺功能不可逆下降、進行性肺瘢痕形成、間質性肺炎等[1]。既往研究表明,肺纖維化的發生和進展是由于不同刺激下肺泡上皮細胞重復損傷后的異常修復[2]。但其確切的發病機制尚不清楚。近年來,一種鐵依賴性、非凋亡調節性的細胞死亡機制被發現并命名為鐵死亡,廣泛參與卒中、腦損傷和腫瘤的發展,然而,鐵是細胞正常生理活動的必需元素,鐵死亡與其他形式的細胞死亡(如凋亡、壞死和自噬)不同,是因為脂質活性氧(reactive oxygen species, ROS)的致命鐵依賴性積累,其原因是細胞內鐵的過度積累導致脂質過氧化升高,從而導致這種類型的細胞死亡[3]。鐵死亡相關基因 GPX4 缺陷小鼠表現出更嚴重的博來霉素誘導的肺纖維化,而轉基因小鼠的這種作用減弱,表明鐵死亡可能在很大程度上促進了肺纖維化[4]。此外,研究表明,肺內鐵負荷過大會導致肺纖維化和肺泡上皮細胞損傷,鐵離子螯合劑如去鐵胺可降低鐵負荷過大引起的肺損傷程度[5]。探究鐵死亡參與肺纖維化的機制有助于發現其潛在治療靶點,因此需進一步的研究來檢測鐵死亡相關基因,包括與肺纖維化有關的基因。因此,本研究旨在探究肺部感染誘導的鐵死亡與肺纖維化的相關性及其機制,為 IFP 的臨床診斷和治療提供新的思路。
1 材料與方法
1.1 生物信息學數據獲取
從基因表達綜合數據庫(Gene Expression Omnibus, GEO)下載 GSE150910 數據集,該數據集涵蓋了 2019 年 1 月—2023 年 12 月 103 個 IPF 肺組織和 103 個正常肺組織的樣本信息,患者信息如表1 所示。該數據集來自公共數據庫,不需要患者同意和倫理委員會的批準。
1.2 差異表達基因(differentially expressed gene, DEG)分析
對于 GSE150910 數據集中的 RNA-seq 初始數據,Reads 計數數據采用每百萬轉錄本(transcripts per million, TPM)進行標準化。TPM 標準化公式如下:TPM=讀取計數×
1.3 鐵死亡相關基因的富集分析
使用 R 3.6.1 軟件中的“GO Plot”包進行基因本體論(Gene Ontology, GO)和京都基因與基因組數據庫(Kyoto Encyclopedia of Genes and Genomes, KEGG)途徑富集分析。GO 分析包括生物過程、細胞成分和分子功能。
1.4 鐵死亡相關 DEG 的蛋白質相互作用(protein-protein interaction, PPI)分析和相關性分析
使用 STRING 在線數據庫和 Cytoscape 3.8.1 軟件分析與鐵死亡相關的 DEG 之間的相互作用,將網絡設置為 STRING 在線數據庫中的截止點(交互評分>0.15)。節點表示基因,邊表示基因之間的相互作用,綠色表示下調基因,藍色表示上調基因。MCODE 是 Cytoscape 的應用程序,用于基因網絡聚類分析,繪制關鍵模塊。利用 R 3.6.1 軟件中的“Corrplot”包(Spearman 相關分析函數)識別差異表達的嗜鐵相關基因之間的相關性,檢驗水準 α=0.05。
1.5 隨機森林法篩選樞紐基因
通過隨機森林算法進一步篩選關鍵模塊中的基因。隨機森林算法是解決生物學研究中各種預測問題的一種有效而流行的分類和回歸方法。對于重要指標的排序,采用隨機森林算法中的平均減少基尼系數來量化哪個指標對分類精度貢獻最大。平均減少基尼系數是一個重要的相關指標,因為其較高的量表明,類別衍生雜質的程度可以通過一個變量來最大程度地降低。使用 GSE150910 開發隨機森林模型,篩選出的前 5 個樞紐基因用于后續研究和實驗驗證。
1.6 樞紐基因的動物實驗驗證
1.6.1 實驗動物及分組
從邯鄲市中心醫院動物實驗中心購買 24 只 8 周齡雄性 C57BL/6 小鼠,體重 20~25 g。將小鼠隨機分為對照組、運動組、博來霉素組、博來霉素+運動組 4 組,每組 6 只。本研究所有程序經邯鄲市中心醫院倫理委員會批準[審查批號:2019 第(22)號],研究內容和過程涉及的動物實驗符合實驗動物福利的倫理要求,符合《動物保護法》及相關規定。動物實驗室設施的環境條件符合中國國家標準《實驗動物與環境設施》(GB 14925—2020)[4],動物飼養管理和動物實驗操作符合邯鄲市中心醫院實驗動物管理規定要求。
1.6.2 博來霉素誘導的肺纖維化動物模型
博來霉素組、博來霉素+運動組小鼠在實驗程序的第 1 天麻醉下(氯胺酮 100 mg/kg 和噻嗪 10 mg/kg)經氣管給藥博來霉素(1.5 U/kg),建立博來霉素誘導的肺纖維化模型[5]。
1.6.3 跑步機運動測試和訓練
如文獻[7]所述,該實驗包含跑步機適應、測試和訓練,小鼠適應跑步機(15 min/d,25° 傾斜,0.2 kg/h)3 d 后,以 0.2 km/h 的起始速度,每 2.5 分鐘增加 0.1 km/h 的速度進行物理測試,實驗以疲勞結束(經過 10 次機械刺激,動物也不能運動)。此后,運動組和博來霉素+運動組小鼠被納入一個為期 4 周、5 次/周、60 min/次的運動訓練計劃,達到最初身體測試中最高速度的 60%。最后一次訓練完成后,將各組小鼠經腹腔內注射氯胺酮/噻嗪(100/10 mg/kg)安樂死,并進行頸椎脫位,收集肺組織用于后續實驗。
1.6.4 實時熒光定量逆轉錄聚合酶鏈反應(real-time quantitative reverse transcription polymerase chain reaction, qRT-PCR)分析
對小鼠肺組織進行 qRT-PCR 分析,驗證鐵死亡標志物前列腺素內過氧化物合成酶 2 和樞紐基因,使用 TRIzol 收集每個肺組織樣本的總 RNA,收集的 RNA 純度用 Quantus 熒光儀測定,首先對總 RNA 進行逆轉錄反應,后將擴增的互補 DNA 樣品與一步 SYBR PrimeScript PLUS RTPCR 試劑盒混合。最后,在 AriaMx HRM 系統上進行反應。該反應的陽性對照為甘油醛-3-磷酸脫氫酶,每個樣品采用比較循環閾值法計算。
1.7 樞紐基因的生存分析
從 GEO 數據庫下載 GSE70866 數據集,該數據集包含來自 212 例 IPF 患者的基因表達譜數據和生存預后信息,用該數據集來驗證樞紐基因的生存預后,用 R 3.6.1 軟件對其進行生存分析。
1.8 統計學方法
使用 GraphPad Prism 8.0 和 R 3.6.1 軟件進行繪圖和統計分析。患者基本信息比較時,計量數據以均數±標準差表示,兩組間比較采用 t 檢驗;計數數據以例數和/或百分數表示,組間比較采用 χ2 檢驗。測序數據使用 R 3.6.1 軟件中的“LIMMA”包進行 DEG 分析,使用“randomForest”包進行樞紐基因篩選。采用單因素方差分析對 4 組小鼠潛在生物標志物的表達量進行統計學分析,并進行 Tukey 多重比較。采用對數秩檢驗分析不同樞紐基因表達水平的 IPF 患者的生存曲線差異。雙側檢驗水準 α=0.05。
2 結果
2.1 鐵死亡相關 DEG
從 GEO 數據庫下載 GSE150910 數據集,得到
2.2 鐵死亡相關 DEG 的富集途徑和分析
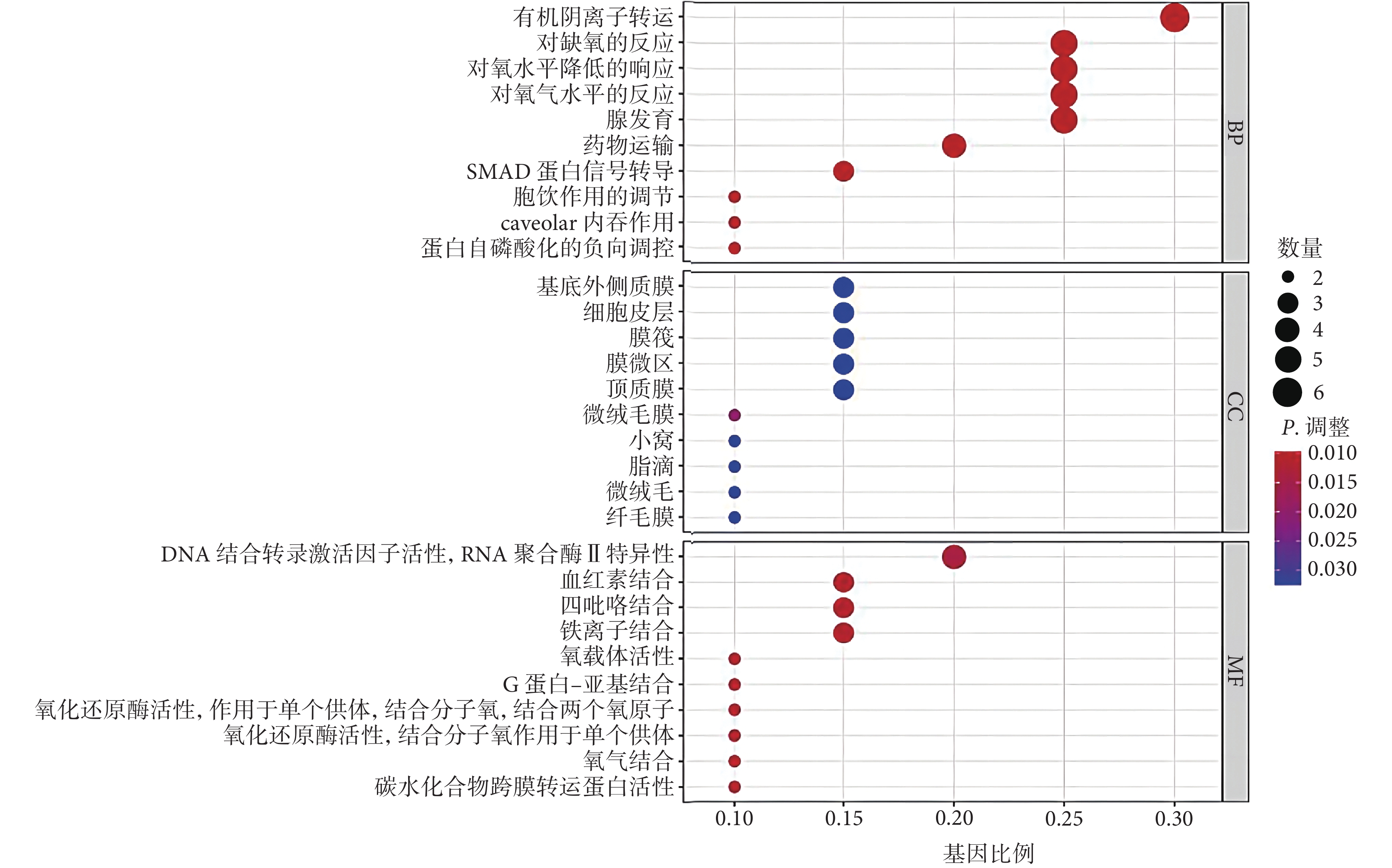
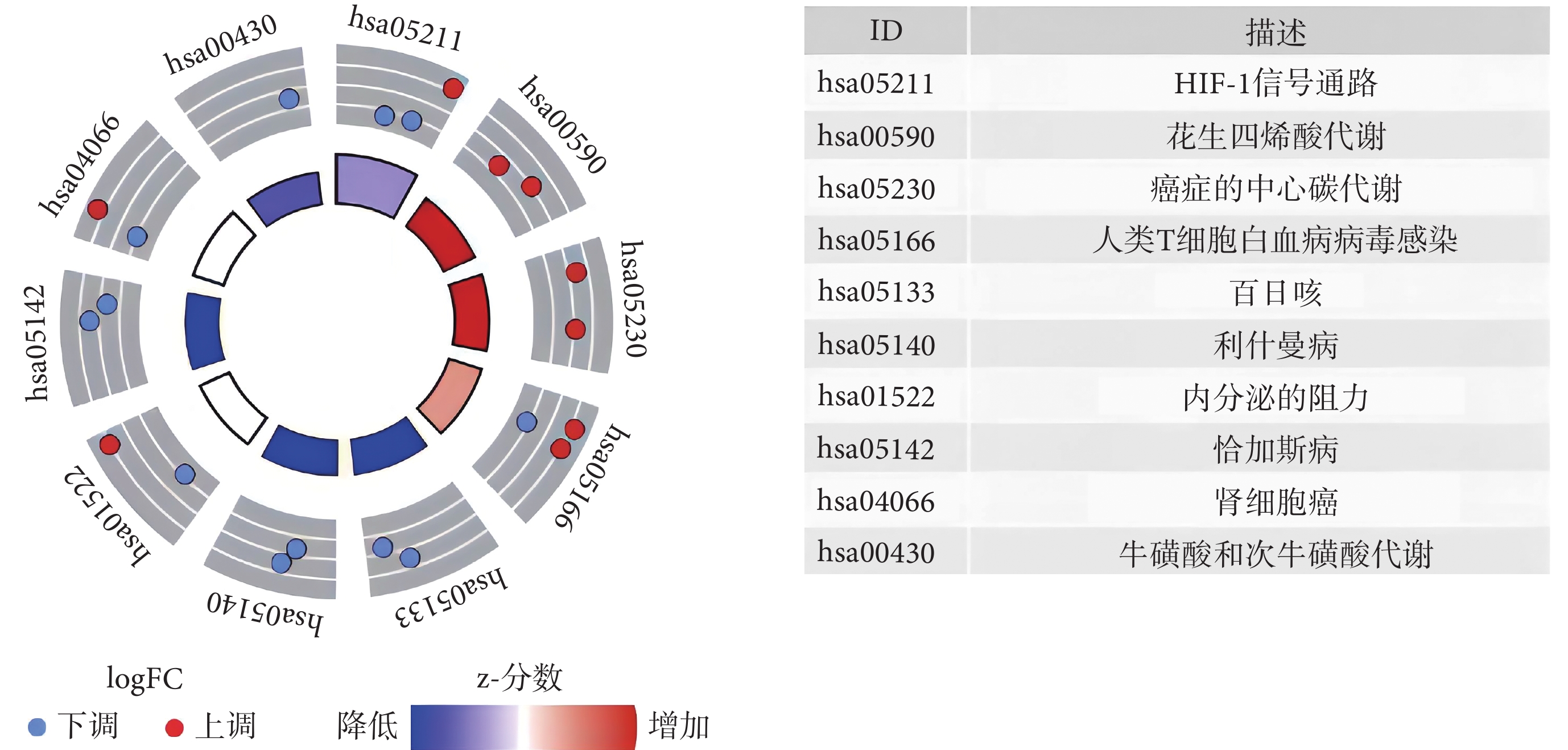
GO 最顯著的富集項包括有機陰離子轉運、對缺氧的反應、對氧水平降低的響應(生物過程),基底外側質膜、細胞皮層、膜筏(細胞成分),DNA-結合轉錄激活因子活性、RNA 聚合酶Ⅱ特異性、血紅素結合、四氟化結合(分子功能),見圖1。KEGG 富集分析結果顯示,與鐵死亡相關的 DEG 主要參與腎細胞癌、花生四烯酸代謝、癌癥中心碳代謝、人 T 細胞白血病病毒 1 感染、百日咳、利什曼病、內分泌抵抗、恰加斯病、缺氧誘導因子-1(hypoxia-inducible factor-1, HIF-1)信號通路、牛磺酸和次牛磺酸代謝(圖2)。
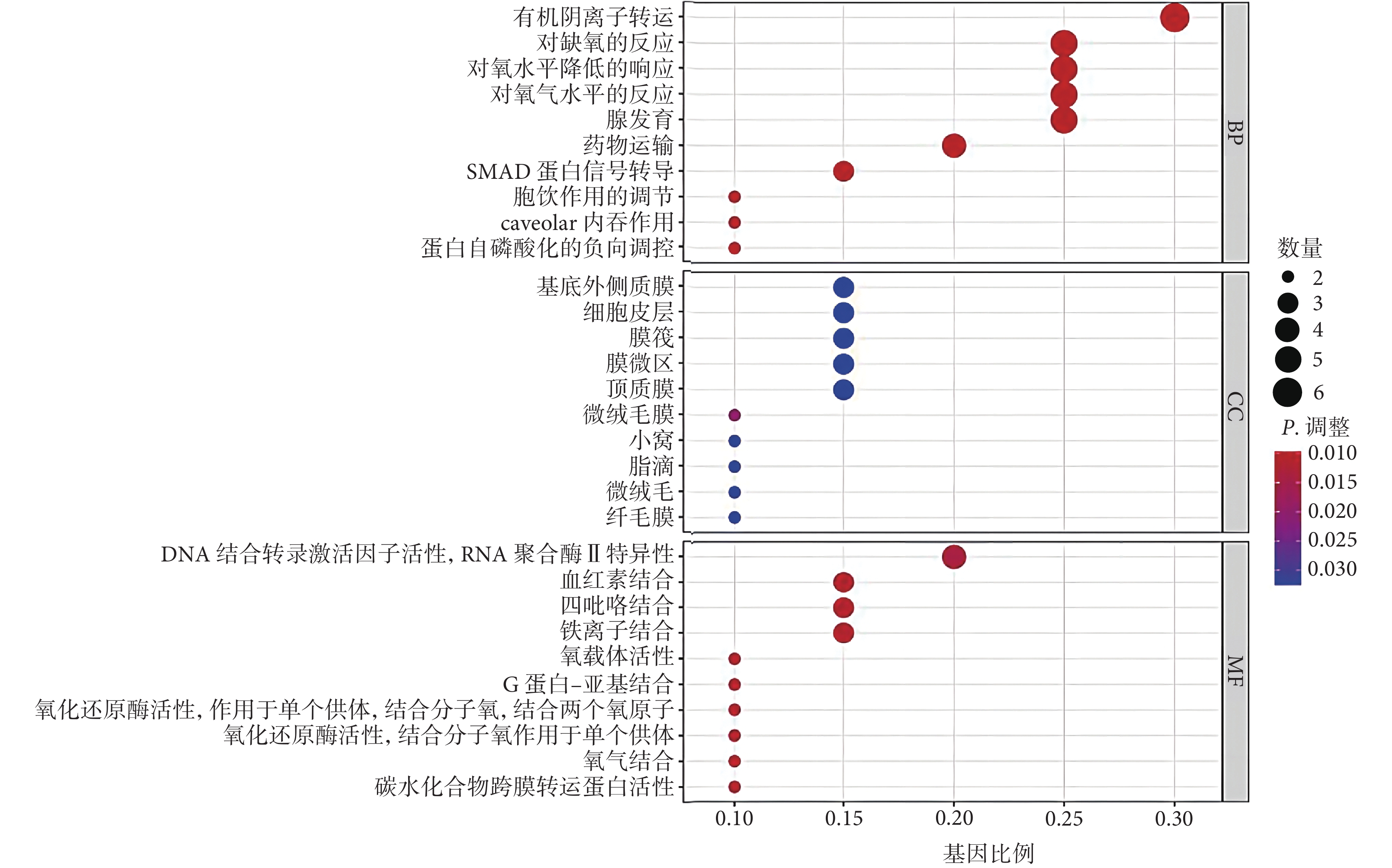 圖1
20 個鐵死亡相關 DEG 的 GO 富集分析
圖1
20 個鐵死亡相關 DEG 的 GO 富集分析
BP:生物過程;CC:細胞成分;MF:分子功能;DEG:差異表達基因;GO:基因本體論
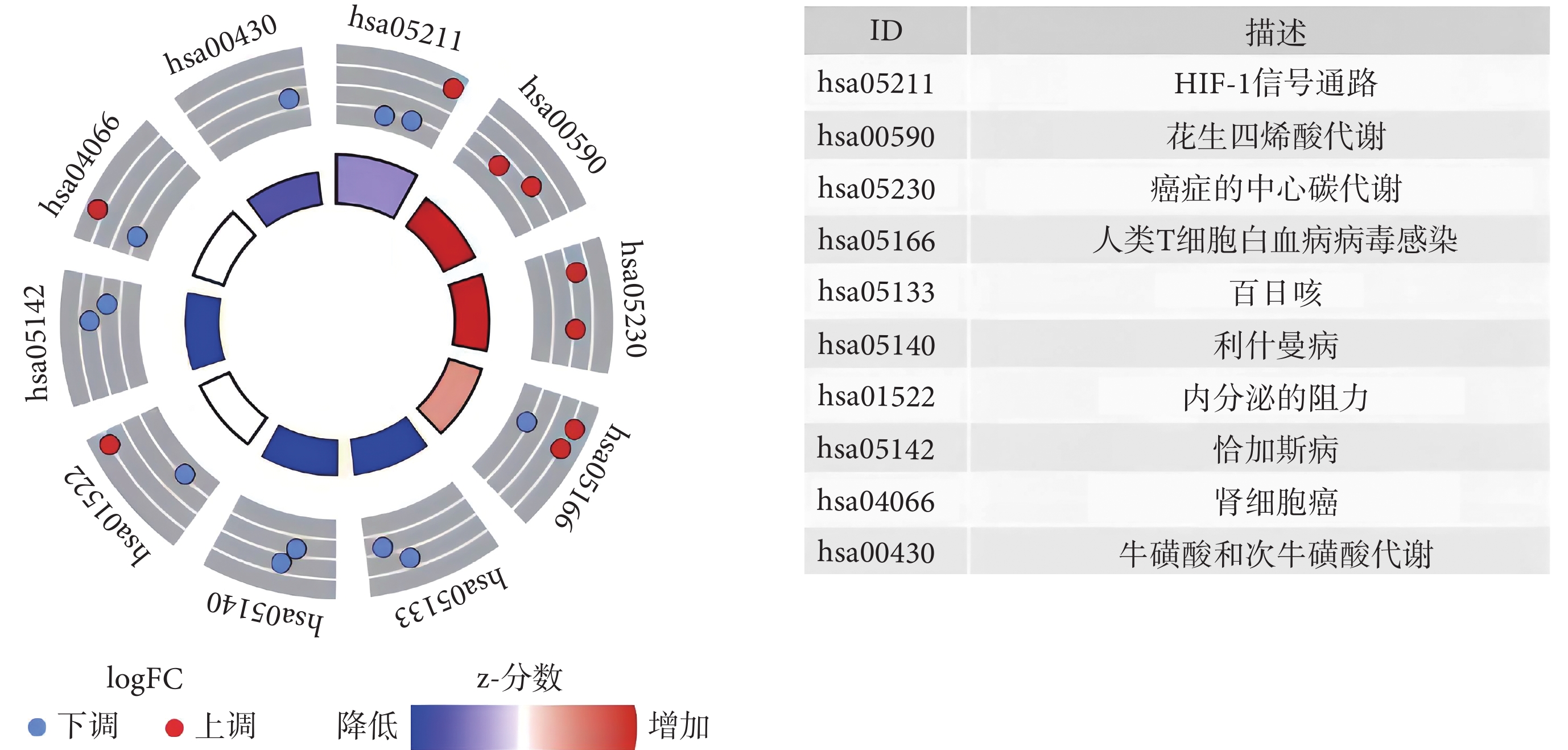 圖2
20 個鐵死亡相關 DEG 的 KEGG 富集分析
圖2
20 個鐵死亡相關 DEG 的 KEGG 富集分析
DEG:差異表達基因;KEGG:京都基因和基因組數據庫;FC:差異倍數;HIF-1:缺氧誘導因子-1
2.3 鐵死亡相關 DEG 的 PPI 網絡和相關性分析
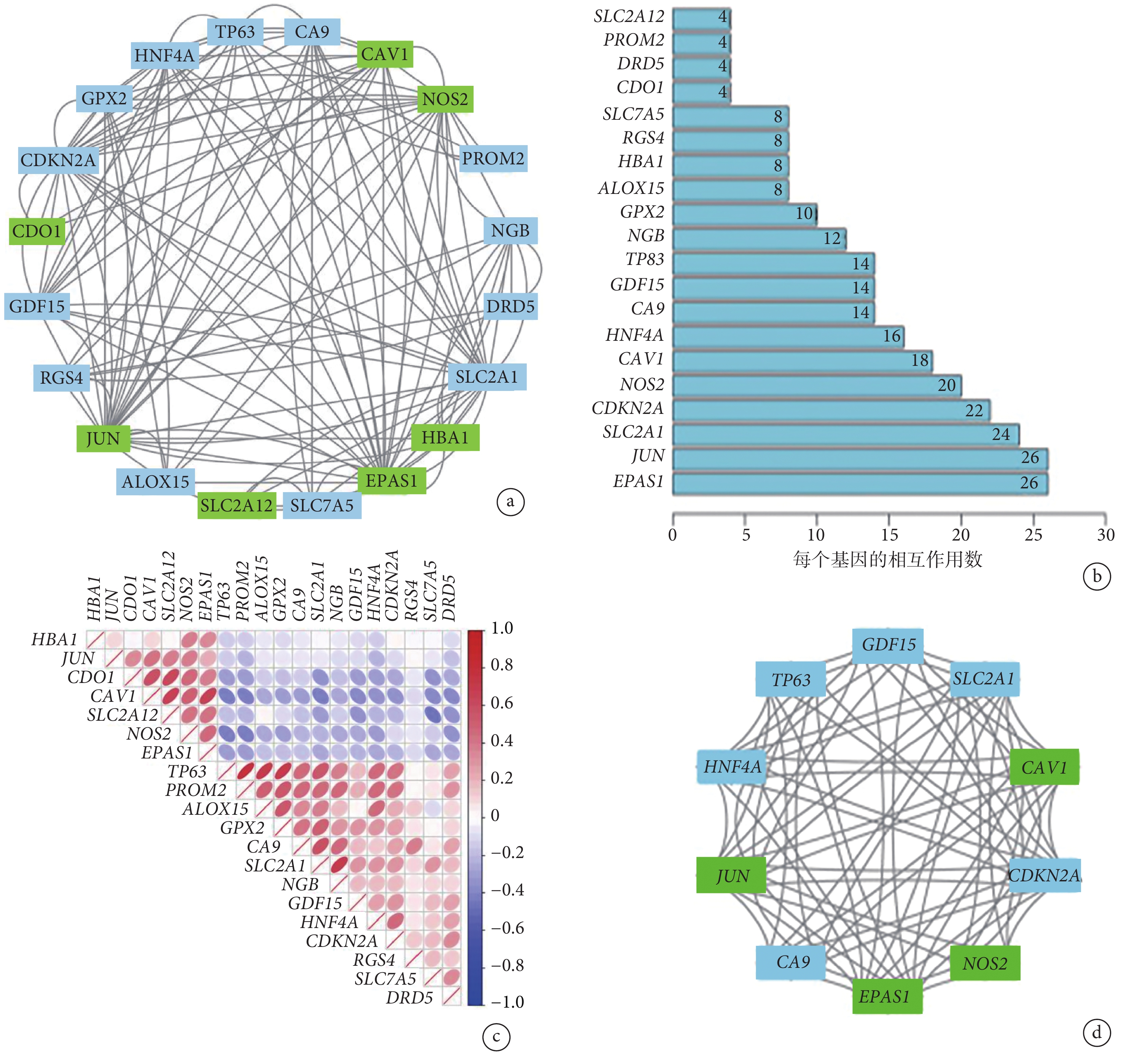
PPI 網絡分析顯示鐵死亡相關 DEG 之間存在相互作用(圖3a)。圖3b 顯示了每個基因的相互作用數量。采用 Spearman 相關分析研究這些基因表達的相關性。來自 GSE150910 數據集的研究結果表明,20 個差異表達的鐵死亡相關基因之間存在相互作用(圖3c)。此外,利用 Cytoscape 的應用程序 MCODE 對基因網絡進行聚類分析。結果,通過繪制關鍵模塊,建立了 4 個切割基因(CAV1、JUN、NOS2、EPAS1)和 6 個凸起基因(GDF15、SLC2A1、CDKN2A、CA9、HNF4A、TP63)(圖3d)。
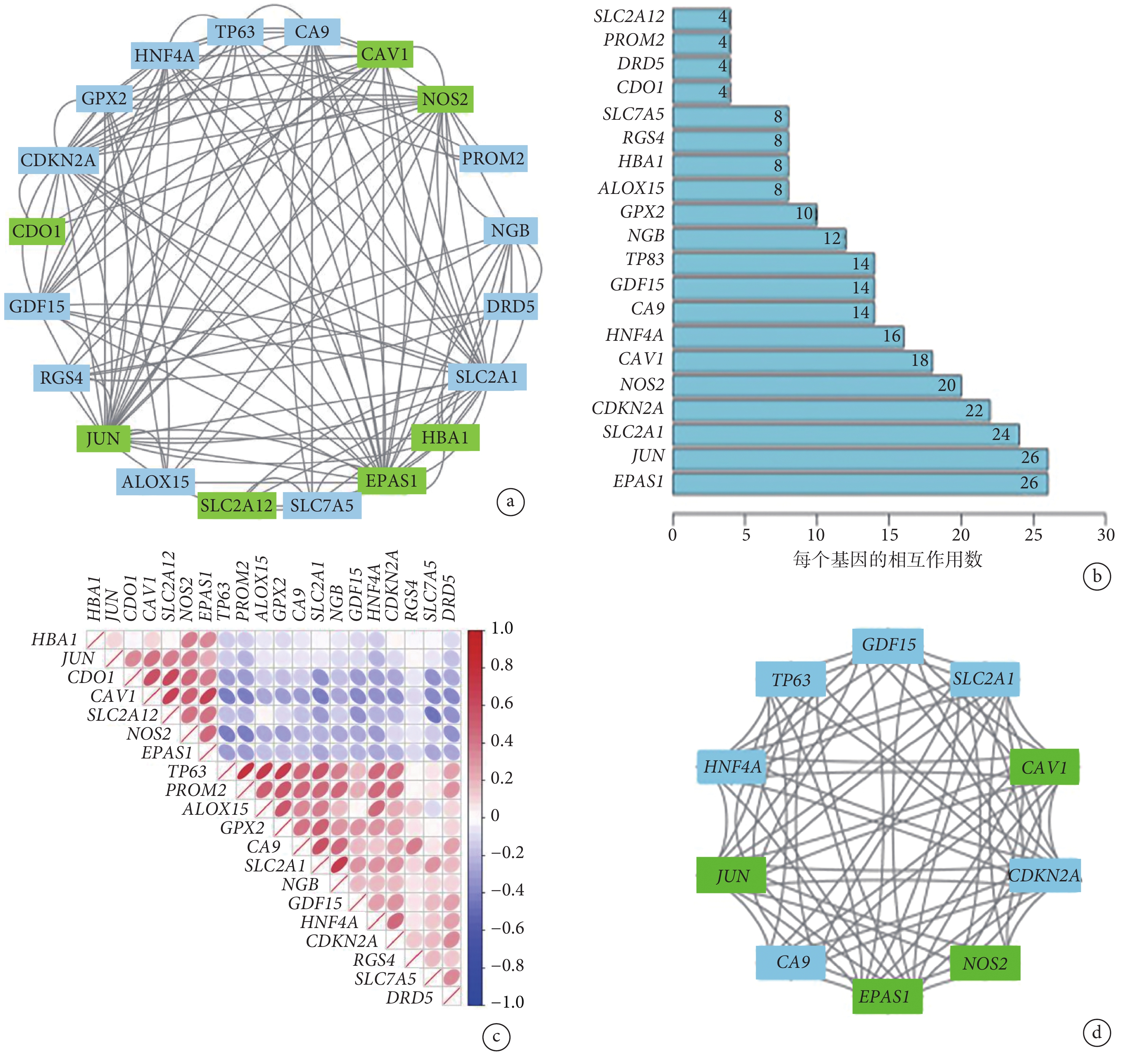 圖3
20 個鐵死亡相關 DEG 的 PPI 網絡分析和相關性分析
圖3
20 個鐵死亡相關 DEG 的 PPI 網絡分析和相關性分析
a. 20 個鐵死亡相關 DEG 中的 PPI 網絡;b. 各 DEG 的相互作用數;c. 20 個鐵死亡相關 DEG 的 Spearman 相關性分析;d. 關鍵模塊由 MCODE 鑒定,用于識別網絡基因聚類。DEG:差異表達基因;PPI:蛋白質相互作用;綠色代表下調基因,藍色代表上調基因
2.4 使用隨機森林分類器識別關鍵 DEG
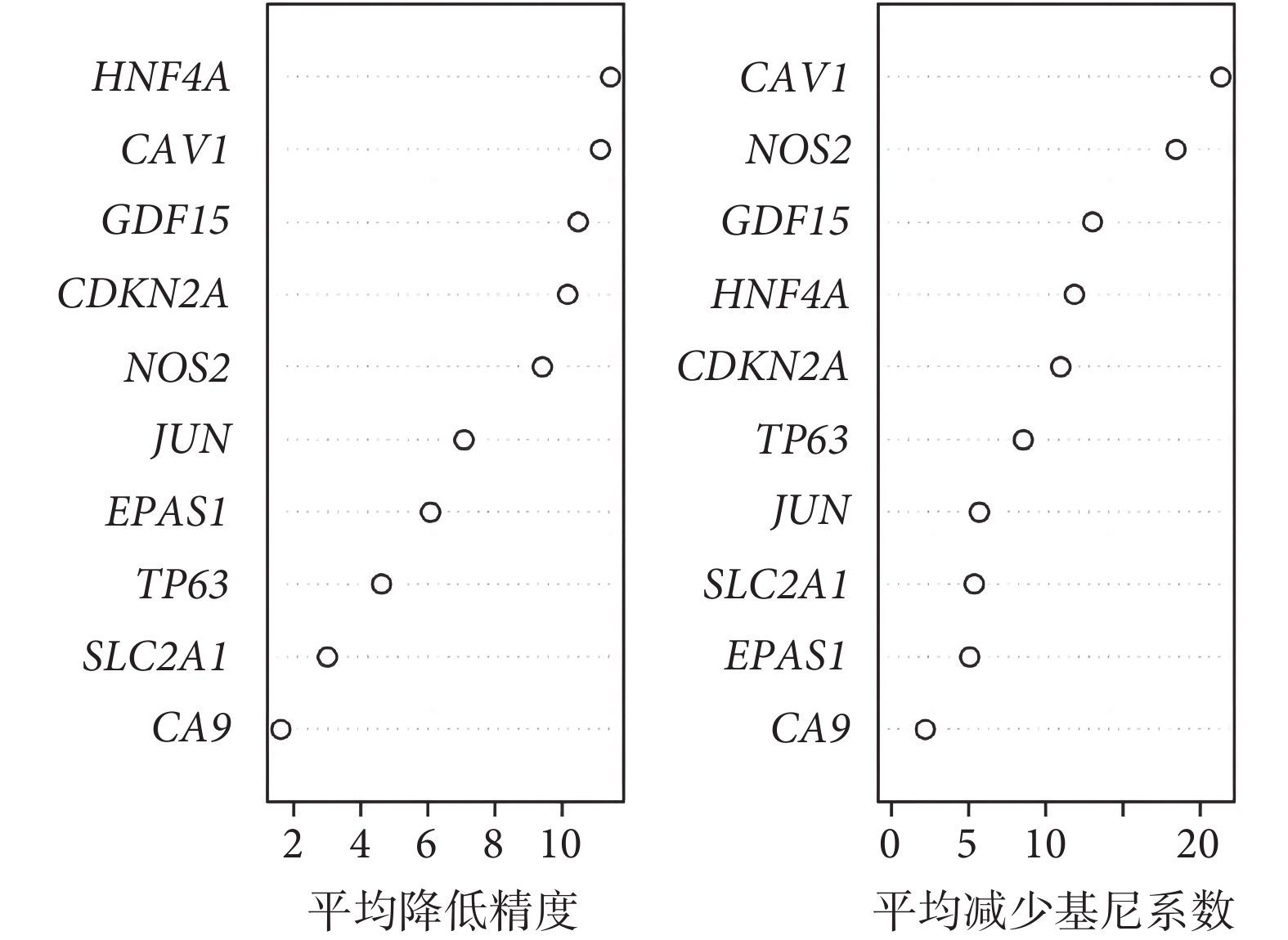
使用隨機森林算法對 MCODE 篩選的 10 個基因進行排名,并選擇前 5 個 DEG(CAV1、NOS2、GDF15、HNF4A、CDKN2A)進行進一步的分析和驗證動物實驗(圖4)。
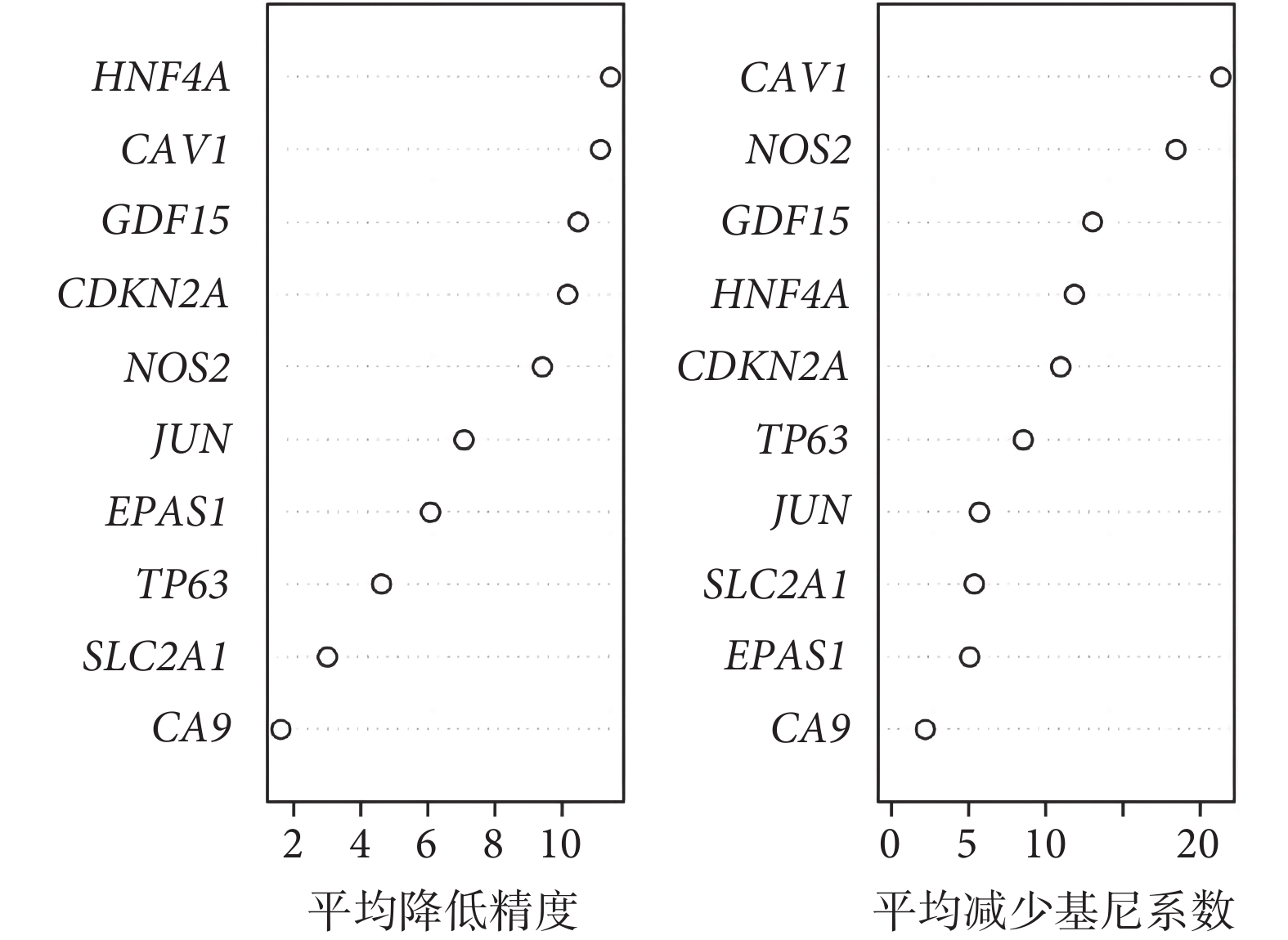 圖4
使用隨機森林分類器鑒定最重要的樞紐基因
圖4
使用隨機森林分類器鑒定最重要的樞紐基因
2.5 qRT-PCR 檢測潛在的生物標志物表達
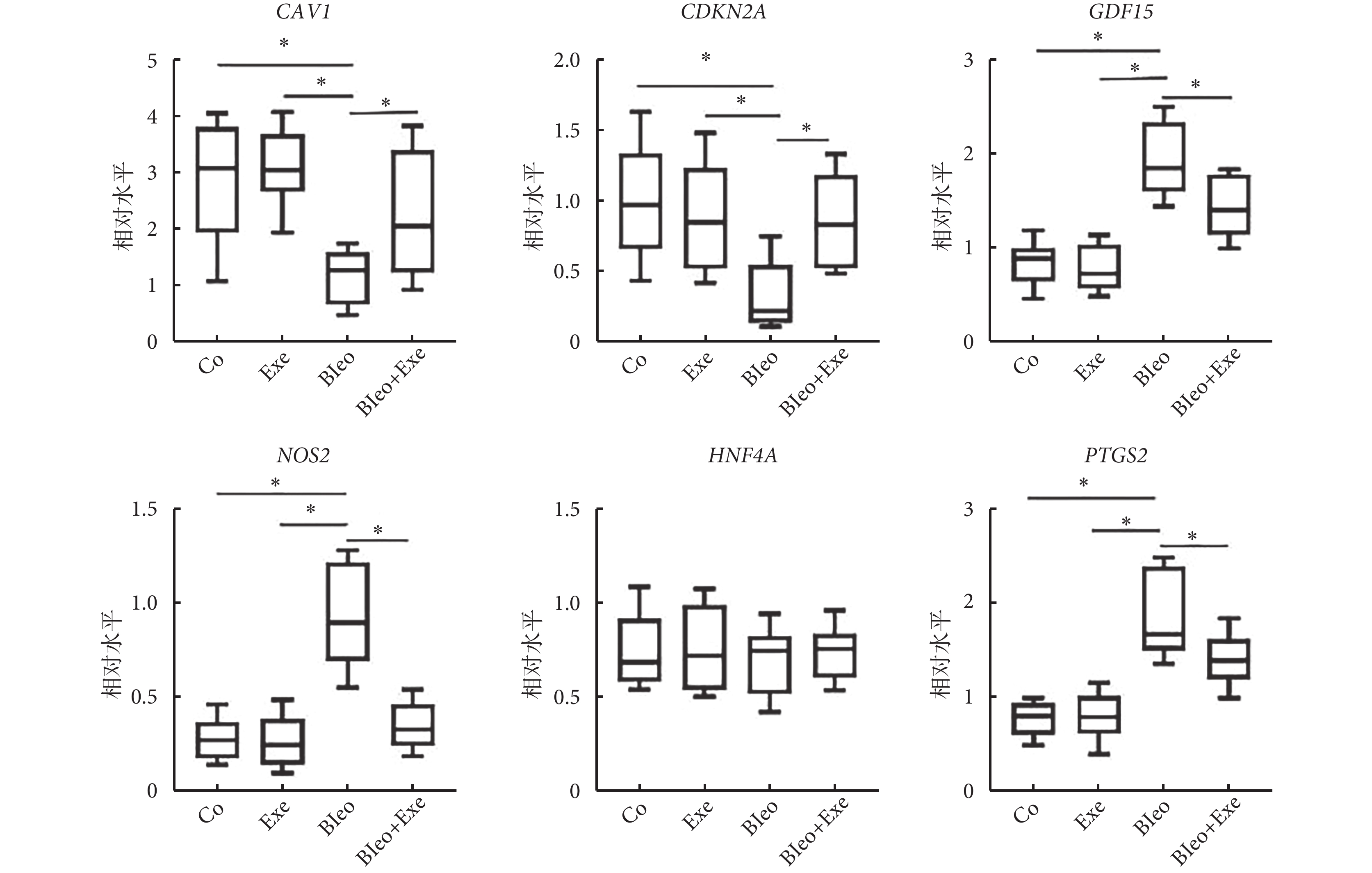
篩選的肺纖維化小鼠模型生物標志物(CAV1、NOS2、GDF15、HNF4A、CDKN2A)和鐵沉標志物 PDGS2 進行 qRT-PCR 驗證。4 組間 HNF4A 的表達水平差異無統計學意義(P>0.05)。運動訓練后,與博來霉素組相比,博來霉素+運動組 CAV1、CDKN2A 表達升高,PTGS2、GDF15、NOS2 表達降低(圖5)。
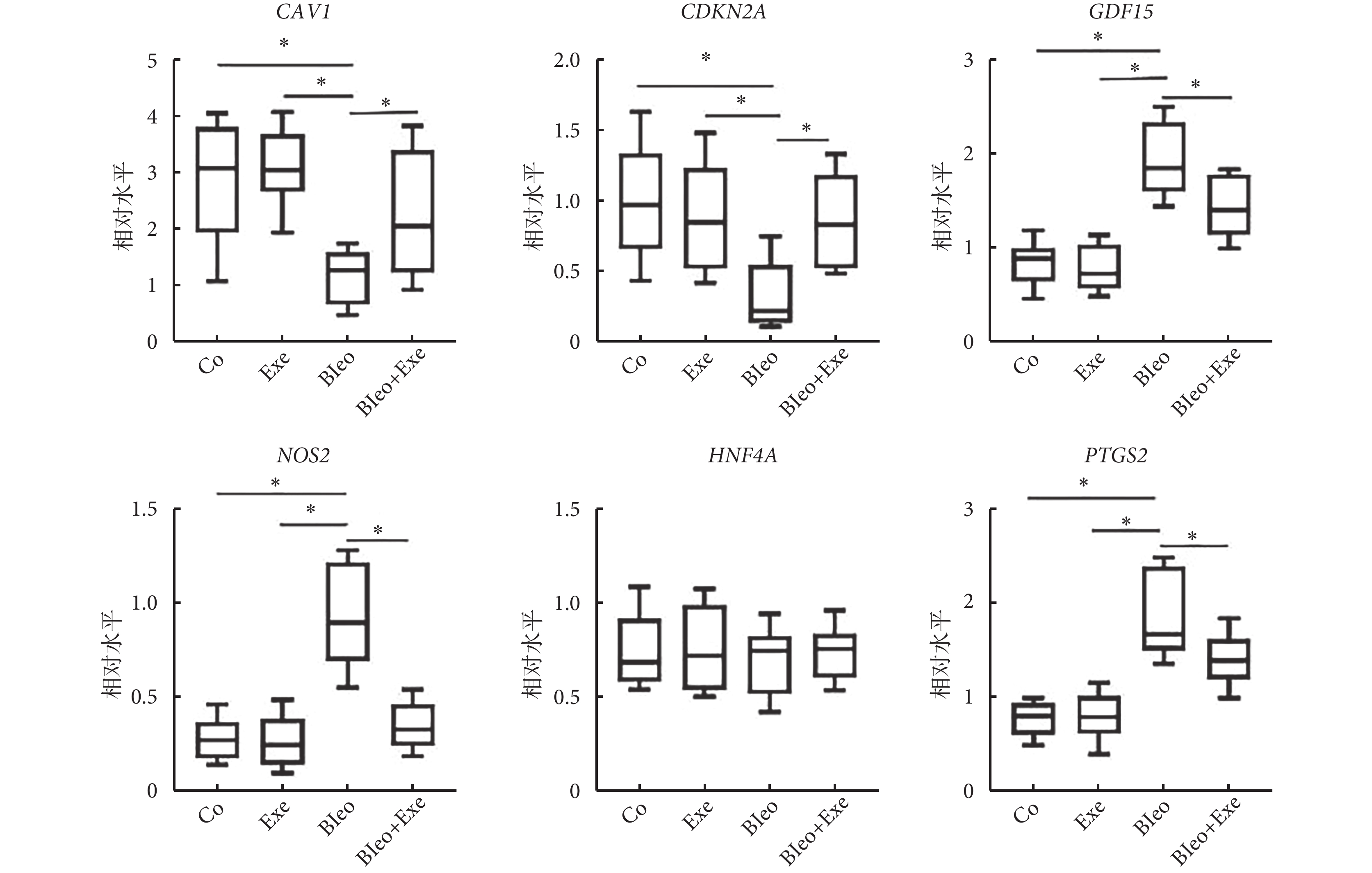 圖5
qRT-PCR 檢測潛在的生物標志物表達
圖5
qRT-PCR 檢測潛在的生物標志物表達
qRT-PCR 檢測肺纖維化小鼠模型生物標志物(
2.6 驗證組中樞紐基因的生存分析
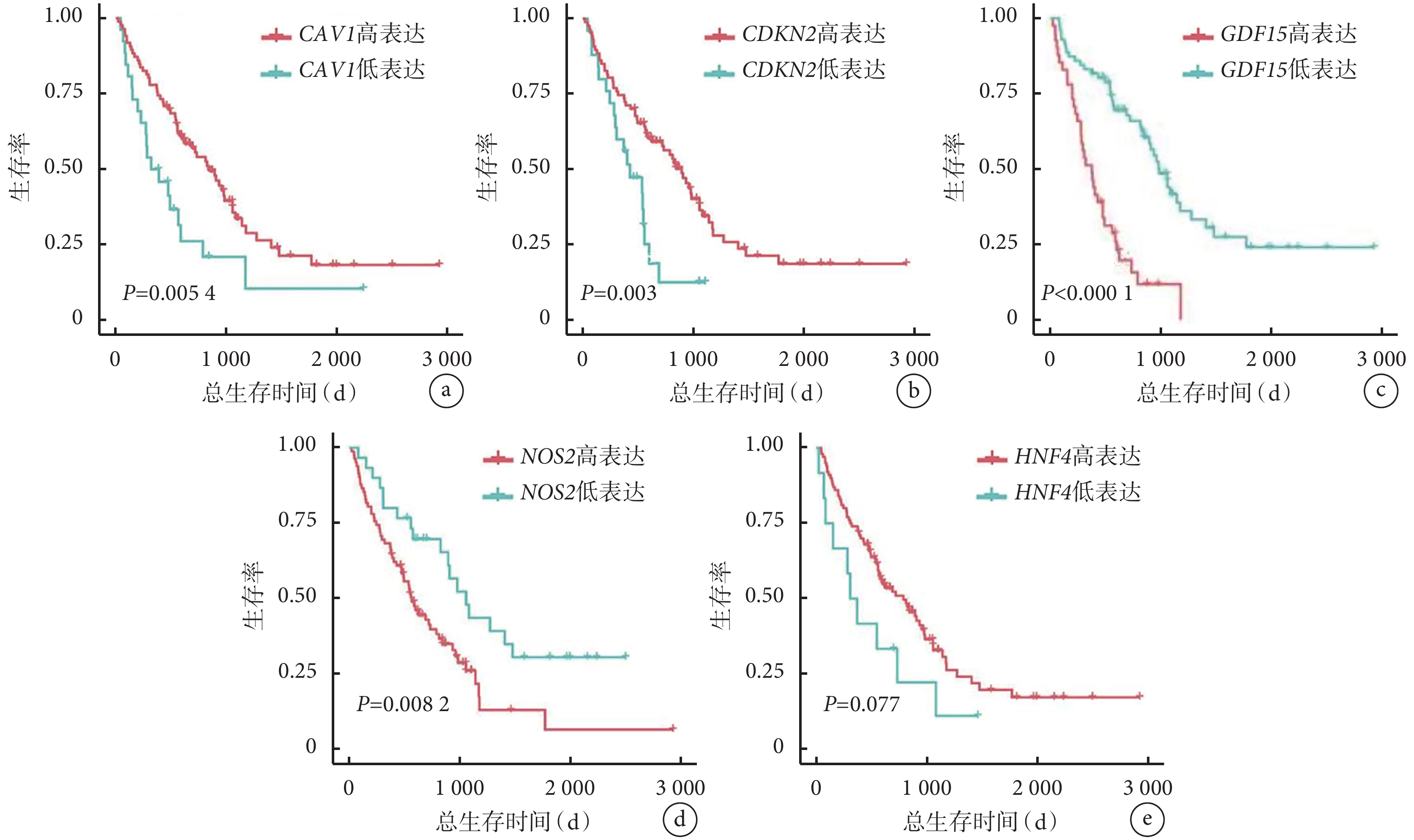
NOS2 和 GDF15 基因表達水平較高的 IPF 患者的總生存率低于這 2 個基因表達水平較低的患者(P<0.05)。此外,與高表達水平的患者相比,IPF 患者中 CAV1 和 CDKN2A 的低表達水平與較差的總生存率相關(P<0.05)。HNF4A 表達水平與 IPF 患者總生存率無關(P>0.05)。見圖6。
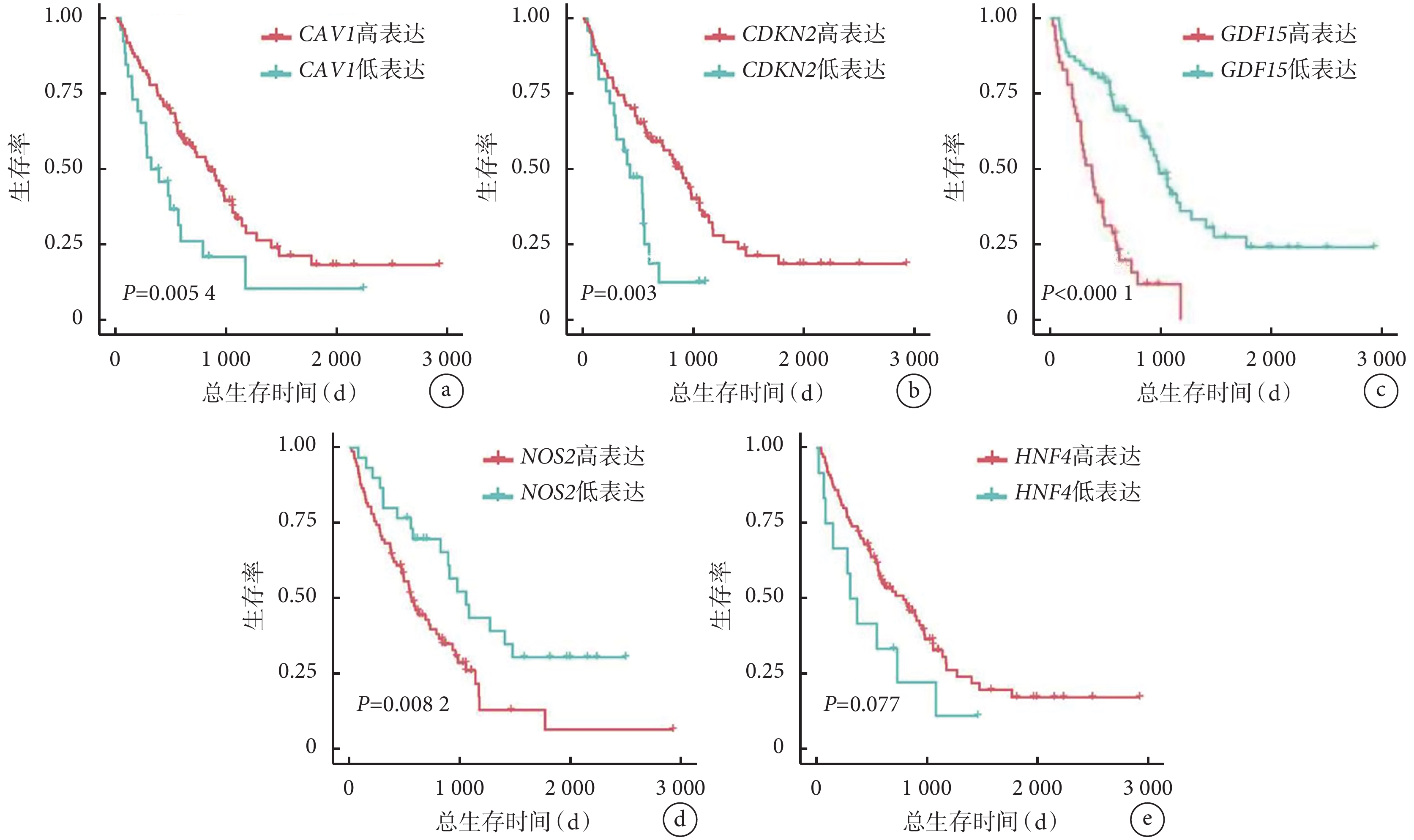 圖6
驗證組 IPF 患者樞紐基因表達水平與總生存時間的 Kaplan-Meier 生存分析
圖6
驗證組 IPF 患者樞紐基因表達水平與總生存時間的 Kaplan-Meier 生存分析
a.
3 討論
本研究確定了 IPF 中鐵死亡的關鍵相關基因,并探討了 IPF 中鐵死亡的潛在發病機制。本研究從 GSE150910 和 FerrDb 數據集的交叉集中共獲得了 20 個基因,包括 13 個上調基因和 7 個下調基因,它們之間存在顯著的相互作用,利用 R 軟件進行富集分析,結果表明這些基因主要參與 HIF-1 信號通路和缺氧反應。因此,本研究采用跑步機訓練來模擬肺纖維化小鼠模型,動物實驗表明,運動可能影響肺纖維化中鐵死亡相關基因的表達,抑制鐵死亡,證實了生物信息學分析的可靠性。
研究表明鐵的過量積累、基于脂質的細胞內 ROS 的過度積累以及最終的脂質氧化是鐵死亡的主要特征,鐵沉會對細胞膜造成損傷,導致細胞死亡,細胞死亡通常是由主要在有氧代謝過程中積累的過量 ROS 引起的氧化應激引起的[8-10]。已有抗氧化劑在肺纖維化中應用,其可能是減少肺纖維化期間鐵死亡,鐵過度積累可以激活 HIF-1 信號通路[11]。這與本研究生物信息學分析一致,本研究富集分析表明 HIF-1 信號通路被激活,誘導肺纖維化后導致鐵死亡或細胞凋亡。
此外,本研究通過 PPI 分析獲得了由 10 個鐵死亡相關基因(CAV1、JUN、NOS2、EPAS1、GDF15、SLC2A1、CDKN2A、CA9、HNF4A、TP63)組成的模塊。對這些基因進行隨機森林分析,篩選出的前 5 個基因分別為 CAV1、NOS2、GDF15、HNF4A、CDKN2A,篩選出的 5 個與鐵死亡相關的基因具有較高的可靠性。
CAV1 是一個膜結合支架蛋白家族,與 CAV2 和 CAV3 蛋白家族相關,CAV1 在肺泡上皮細胞、內皮細胞、成纖維細胞、平滑肌細胞等肺組織中廣泛表達,許多研究也討論了 IPF 患者肺組織中 CAV1 表達明顯降低[12],這與本研究生物信息學分析結果一致。原因是正常表達 CAV1 可以抑制轉化生長因子(transforming growth factor, TGF)-β 的過表達,CAV1 表達降低會減弱 TGF-β 的抑制作用,從而激活 TGF-β 信號通路,導致細胞外基質大量生成,導致肺纖維化的發生[13]。本研究中的肺纖維化小鼠模型進一步顯示了在博來霉素誘導的肺纖維化組織中,CAV1 mRNA 的表達水平明顯低于正常對照組;總體生存分析顯示,CAV1 高表達患者的生存時間更長。CAV1 是纖維素的主要成分,調節細胞信號傳導和內吞作用,過表達 CAV1 可以減少中性粒細胞和單核/巨噬細胞的浸潤,防止博來霉素誘導的肺纖維化,這可能與其在炎癥活動中的關鍵調節作用有關[14-15]。本研究動物實驗表明,在肺纖維化小鼠中進行跑步機運動時,CAV1 mRNA 的表達增加。相關研究表明,在博來霉素誘導的肺纖維化模型中,跑步機運動可改善肺部和全身炎癥[14],這可能與本研究中跑步機增加 CAV1 有關。
CDKN2A 被稱為周期依賴性激酶抑制劑基因,屬于周期依賴性激酶抑制劑家族,是重要的腫瘤抑制基因,IPF 患者外周血 CDKN2A mRNA 表達水平較對照組降低,可能與 P53 信號通路激活有關[16]。在本實驗中,肺纖維化小鼠肺組織中 CDKN2A mRNA的表達水平低于正常對照組。FERDB 在線工具預測 CDKN2A 與鐵死亡相關,并將其歸類為鐵死亡驅動因子,CDKN2A 是否是 IPF 中鐵死亡的驅動因素尚未明確。因此,本研究結果表明 CDKN2A 影響肺纖維化的鐵死亡,其機制值得進一步研究。
NOS2 是氧化代謝的關鍵基因。既往研究報道,在肺纖維化病理生理條件下,存在異常高濃度的一氧化氮,這可能與誘導型一氧化氮合酶活性升高有關,過量的一氧化氮產生可能有助于纖維化的發展[17-18]。GDF15 是一種內分泌激素,是 TGF-β 超家族的成員,可促進腫瘤細胞的鐵死亡;GDF15 是一種上皮來源的分泌蛋白,可能是一種有用的上皮應激生物標志物,因為其表達增加與 IPF 患者預后較差相關[19],這與本研究當前生物信息學分析的結果一致。HNF4A 是一種核轉錄因子,以同型二聚體的形式與 DNA 結合,主要與肝臟纖維化的發生有關,敲除 HNF4A 可緩解肝纖維化[20],然而沒有關于 HNF4A 與肺纖維化相關性的研究報道。同樣,本研究生物信息學分析的動物實驗和生存分析表明,HNF4A 與肺纖維化無關聯。本研究生物信息學分析數據表明,IPF 組織中 NOS2 和 CAV1 下調,CDKN2A 和 GDF15 上調;但在博來霉素誘導的小鼠 IPF 組織中,NOS2 上調,CDKN2A 下調。實驗結果與生物信息學分析結果的不一致可能是由于以下原因:① 用于生物信息學分析的數據來自 IPF 患者,而實驗數據來自博來霉素誘導的小鼠,雖然人和小鼠具有很高的基因同源性,但由于物種不同,結果可能存在一定的偏差;② 基因擴增具有很大的時間和空間特異性,從而導致生物信息學分析與實驗結果不一致。
本研究有以下局限性:首先,從 IPF 患者肺組織和正常肺組織中獲取生物信息學結果,由于實驗條件和醫院規模的限制,臨床組織樣本收集不足進行驗證。此外,動物模型中不同鐵死亡基因表達水平的驗證,缺乏樞紐基因在肺纖維化細胞模型中的潛在研究。
綜上所述,通過生物信息學分析,本研究確定了 20 個肺纖維化中與鐵死亡相關的潛在基因。CAV1、NOS2、GDF15、CDKN2A 可能通過調控鐵死亡影響肺纖維化的發生。肺纖維化誘導后的有氧運動訓練可減輕肺鐵死亡,可能有助于肺纖維化的治療。
利益沖突:所有作者聲明不存在利益沖突。
特發性肺纖維化(idiopathic pulmonary fibrosis, IPF)是一種進行性、慢性、不可逆的肺部疾病,其癥狀包括肺功能不可逆下降、進行性肺瘢痕形成、間質性肺炎等[1]。既往研究表明,肺纖維化的發生和進展是由于不同刺激下肺泡上皮細胞重復損傷后的異常修復[2]。但其確切的發病機制尚不清楚。近年來,一種鐵依賴性、非凋亡調節性的細胞死亡機制被發現并命名為鐵死亡,廣泛參與卒中、腦損傷和腫瘤的發展,然而,鐵是細胞正常生理活動的必需元素,鐵死亡與其他形式的細胞死亡(如凋亡、壞死和自噬)不同,是因為脂質活性氧(reactive oxygen species, ROS)的致命鐵依賴性積累,其原因是細胞內鐵的過度積累導致脂質過氧化升高,從而導致這種類型的細胞死亡[3]。鐵死亡相關基因 GPX4 缺陷小鼠表現出更嚴重的博來霉素誘導的肺纖維化,而轉基因小鼠的這種作用減弱,表明鐵死亡可能在很大程度上促進了肺纖維化[4]。此外,研究表明,肺內鐵負荷過大會導致肺纖維化和肺泡上皮細胞損傷,鐵離子螯合劑如去鐵胺可降低鐵負荷過大引起的肺損傷程度[5]。探究鐵死亡參與肺纖維化的機制有助于發現其潛在治療靶點,因此需進一步的研究來檢測鐵死亡相關基因,包括與肺纖維化有關的基因。因此,本研究旨在探究肺部感染誘導的鐵死亡與肺纖維化的相關性及其機制,為 IFP 的臨床診斷和治療提供新的思路。
1 材料與方法
1.1 生物信息學數據獲取
從基因表達綜合數據庫(Gene Expression Omnibus, GEO)下載 GSE150910 數據集,該數據集涵蓋了 2019 年 1 月—2023 年 12 月 103 個 IPF 肺組織和 103 個正常肺組織的樣本信息,患者信息如表1 所示。該數據集來自公共數據庫,不需要患者同意和倫理委員會的批準。
1.2 差異表達基因(differentially expressed gene, DEG)分析
對于 GSE150910 數據集中的 RNA-seq 初始數據,Reads 計數數據采用每百萬轉錄本(transcripts per million, TPM)進行標準化。TPM 標準化公式如下:TPM=讀取計數×
1.3 鐵死亡相關基因的富集分析
使用 R 3.6.1 軟件中的“GO Plot”包進行基因本體論(Gene Ontology, GO)和京都基因與基因組數據庫(Kyoto Encyclopedia of Genes and Genomes, KEGG)途徑富集分析。GO 分析包括生物過程、細胞成分和分子功能。
1.4 鐵死亡相關 DEG 的蛋白質相互作用(protein-protein interaction, PPI)分析和相關性分析
使用 STRING 在線數據庫和 Cytoscape 3.8.1 軟件分析與鐵死亡相關的 DEG 之間的相互作用,將網絡設置為 STRING 在線數據庫中的截止點(交互評分>0.15)。節點表示基因,邊表示基因之間的相互作用,綠色表示下調基因,藍色表示上調基因。MCODE 是 Cytoscape 的應用程序,用于基因網絡聚類分析,繪制關鍵模塊。利用 R 3.6.1 軟件中的“Corrplot”包(Spearman 相關分析函數)識別差異表達的嗜鐵相關基因之間的相關性,檢驗水準 α=0.05。
1.5 隨機森林法篩選樞紐基因
通過隨機森林算法進一步篩選關鍵模塊中的基因。隨機森林算法是解決生物學研究中各種預測問題的一種有效而流行的分類和回歸方法。對于重要指標的排序,采用隨機森林算法中的平均減少基尼系數來量化哪個指標對分類精度貢獻最大。平均減少基尼系數是一個重要的相關指標,因為其較高的量表明,類別衍生雜質的程度可以通過一個變量來最大程度地降低。使用 GSE150910 開發隨機森林模型,篩選出的前 5 個樞紐基因用于后續研究和實驗驗證。
1.6 樞紐基因的動物實驗驗證
1.6.1 實驗動物及分組
從邯鄲市中心醫院動物實驗中心購買 24 只 8 周齡雄性 C57BL/6 小鼠,體重 20~25 g。將小鼠隨機分為對照組、運動組、博來霉素組、博來霉素+運動組 4 組,每組 6 只。本研究所有程序經邯鄲市中心醫院倫理委員會批準[審查批號:2019 第(22)號],研究內容和過程涉及的動物實驗符合實驗動物福利的倫理要求,符合《動物保護法》及相關規定。動物實驗室設施的環境條件符合中國國家標準《實驗動物與環境設施》(GB 14925—2020)[4],動物飼養管理和動物實驗操作符合邯鄲市中心醫院實驗動物管理規定要求。
1.6.2 博來霉素誘導的肺纖維化動物模型
博來霉素組、博來霉素+運動組小鼠在實驗程序的第 1 天麻醉下(氯胺酮 100 mg/kg 和噻嗪 10 mg/kg)經氣管給藥博來霉素(1.5 U/kg),建立博來霉素誘導的肺纖維化模型[5]。
1.6.3 跑步機運動測試和訓練
如文獻[7]所述,該實驗包含跑步機適應、測試和訓練,小鼠適應跑步機(15 min/d,25° 傾斜,0.2 kg/h)3 d 后,以 0.2 km/h 的起始速度,每 2.5 分鐘增加 0.1 km/h 的速度進行物理測試,實驗以疲勞結束(經過 10 次機械刺激,動物也不能運動)。此后,運動組和博來霉素+運動組小鼠被納入一個為期 4 周、5 次/周、60 min/次的運動訓練計劃,達到最初身體測試中最高速度的 60%。最后一次訓練完成后,將各組小鼠經腹腔內注射氯胺酮/噻嗪(100/10 mg/kg)安樂死,并進行頸椎脫位,收集肺組織用于后續實驗。
1.6.4 實時熒光定量逆轉錄聚合酶鏈反應(real-time quantitative reverse transcription polymerase chain reaction, qRT-PCR)分析
對小鼠肺組織進行 qRT-PCR 分析,驗證鐵死亡標志物前列腺素內過氧化物合成酶 2 和樞紐基因,使用 TRIzol 收集每個肺組織樣本的總 RNA,收集的 RNA 純度用 Quantus 熒光儀測定,首先對總 RNA 進行逆轉錄反應,后將擴增的互補 DNA 樣品與一步 SYBR PrimeScript PLUS RTPCR 試劑盒混合。最后,在 AriaMx HRM 系統上進行反應。該反應的陽性對照為甘油醛-3-磷酸脫氫酶,每個樣品采用比較循環閾值法計算。
1.7 樞紐基因的生存分析
從 GEO 數據庫下載 GSE70866 數據集,該數據集包含來自 212 例 IPF 患者的基因表達譜數據和生存預后信息,用該數據集來驗證樞紐基因的生存預后,用 R 3.6.1 軟件對其進行生存分析。
1.8 統計學方法
使用 GraphPad Prism 8.0 和 R 3.6.1 軟件進行繪圖和統計分析。患者基本信息比較時,計量數據以均數±標準差表示,兩組間比較采用 t 檢驗;計數數據以例數和/或百分數表示,組間比較采用 χ2 檢驗。測序數據使用 R 3.6.1 軟件中的“LIMMA”包進行 DEG 分析,使用“randomForest”包進行樞紐基因篩選。采用單因素方差分析對 4 組小鼠潛在生物標志物的表達量進行統計學分析,并進行 Tukey 多重比較。采用對數秩檢驗分析不同樞紐基因表達水平的 IPF 患者的生存曲線差異。雙側檢驗水準 α=0.05。
2 結果
2.1 鐵死亡相關 DEG
從 GEO 數據庫下載 GSE150910 數據集,得到
2.2 鐵死亡相關 DEG 的富集途徑和分析
GO 最顯著的富集項包括有機陰離子轉運、對缺氧的反應、對氧水平降低的響應(生物過程),基底外側質膜、細胞皮層、膜筏(細胞成分),DNA-結合轉錄激活因子活性、RNA 聚合酶Ⅱ特異性、血紅素結合、四氟化結合(分子功能),見圖1。KEGG 富集分析結果顯示,與鐵死亡相關的 DEG 主要參與腎細胞癌、花生四烯酸代謝、癌癥中心碳代謝、人 T 細胞白血病病毒 1 感染、百日咳、利什曼病、內分泌抵抗、恰加斯病、缺氧誘導因子-1(hypoxia-inducible factor-1, HIF-1)信號通路、牛磺酸和次牛磺酸代謝(圖2)。
圖1
20 個鐵死亡相關 DEG 的 GO 富集分析
BP:生物過程;CC:細胞成分;MF:分子功能;DEG:差異表達基因;GO:基因本體論
圖2
20 個鐵死亡相關 DEG 的 KEGG 富集分析
DEG:差異表達基因;KEGG:京都基因和基因組數據庫;FC:差異倍數;HIF-1:缺氧誘導因子-1
2.3 鐵死亡相關 DEG 的 PPI 網絡和相關性分析
PPI 網絡分析顯示鐵死亡相關 DEG 之間存在相互作用(圖3a)。圖3b 顯示了每個基因的相互作用數量。采用 Spearman 相關分析研究這些基因表達的相關性。來自 GSE150910 數據集的研究結果表明,20 個差異表達的鐵死亡相關基因之間存在相互作用(圖3c)。此外,利用 Cytoscape 的應用程序 MCODE 對基因網絡進行聚類分析。結果,通過繪制關鍵模塊,建立了 4 個切割基因(CAV1、JUN、NOS2、EPAS1)和 6 個凸起基因(GDF15、SLC2A1、CDKN2A、CA9、HNF4A、TP63)(圖3d)。
圖3
20 個鐵死亡相關 DEG 的 PPI 網絡分析和相關性分析
a. 20 個鐵死亡相關 DEG 中的 PPI 網絡;b. 各 DEG 的相互作用數;c. 20 個鐵死亡相關 DEG 的 Spearman 相關性分析;d. 關鍵模塊由 MCODE 鑒定,用于識別網絡基因聚類。DEG:差異表達基因;PPI:蛋白質相互作用;綠色代表下調基因,藍色代表上調基因
2.4 使用隨機森林分類器識別關鍵 DEG
使用隨機森林算法對 MCODE 篩選的 10 個基因進行排名,并選擇前 5 個 DEG(CAV1、NOS2、GDF15、HNF4A、CDKN2A)進行進一步的分析和驗證動物實驗(圖4)。
圖4
使用隨機森林分類器鑒定最重要的樞紐基因
2.5 qRT-PCR 檢測潛在的生物標志物表達
篩選的肺纖維化小鼠模型生物標志物(CAV1、NOS2、GDF15、HNF4A、CDKN2A)和鐵沉標志物 PDGS2 進行 qRT-PCR 驗證。4 組間 HNF4A 的表達水平差異無統計學意義(P>0.05)。運動訓練后,與博來霉素組相比,博來霉素+運動組 CAV1、CDKN2A 表達升高,PTGS2、GDF15、NOS2 表達降低(圖5)。
圖5
qRT-PCR 檢測潛在的生物標志物表達
qRT-PCR 檢測肺纖維化小鼠模型生物標志物(
2.6 驗證組中樞紐基因的生存分析
NOS2 和 GDF15 基因表達水平較高的 IPF 患者的總生存率低于這 2 個基因表達水平較低的患者(P<0.05)。此外,與高表達水平的患者相比,IPF 患者中 CAV1 和 CDKN2A 的低表達水平與較差的總生存率相關(P<0.05)。HNF4A 表達水平與 IPF 患者總生存率無關(P>0.05)。見圖6。
圖6
驗證組 IPF 患者樞紐基因表達水平與總生存時間的 Kaplan-Meier 生存分析
a.
3 討論
本研究確定了 IPF 中鐵死亡的關鍵相關基因,并探討了 IPF 中鐵死亡的潛在發病機制。本研究從 GSE150910 和 FerrDb 數據集的交叉集中共獲得了 20 個基因,包括 13 個上調基因和 7 個下調基因,它們之間存在顯著的相互作用,利用 R 軟件進行富集分析,結果表明這些基因主要參與 HIF-1 信號通路和缺氧反應。因此,本研究采用跑步機訓練來模擬肺纖維化小鼠模型,動物實驗表明,運動可能影響肺纖維化中鐵死亡相關基因的表達,抑制鐵死亡,證實了生物信息學分析的可靠性。
研究表明鐵的過量積累、基于脂質的細胞內 ROS 的過度積累以及最終的脂質氧化是鐵死亡的主要特征,鐵沉會對細胞膜造成損傷,導致細胞死亡,細胞死亡通常是由主要在有氧代謝過程中積累的過量 ROS 引起的氧化應激引起的[8-10]。已有抗氧化劑在肺纖維化中應用,其可能是減少肺纖維化期間鐵死亡,鐵過度積累可以激活 HIF-1 信號通路[11]。這與本研究生物信息學分析一致,本研究富集分析表明 HIF-1 信號通路被激活,誘導肺纖維化后導致鐵死亡或細胞凋亡。
此外,本研究通過 PPI 分析獲得了由 10 個鐵死亡相關基因(CAV1、JUN、NOS2、EPAS1、GDF15、SLC2A1、CDKN2A、CA9、HNF4A、TP63)組成的模塊。對這些基因進行隨機森林分析,篩選出的前 5 個基因分別為 CAV1、NOS2、GDF15、HNF4A、CDKN2A,篩選出的 5 個與鐵死亡相關的基因具有較高的可靠性。
CAV1 是一個膜結合支架蛋白家族,與 CAV2 和 CAV3 蛋白家族相關,CAV1 在肺泡上皮細胞、內皮細胞、成纖維細胞、平滑肌細胞等肺組織中廣泛表達,許多研究也討論了 IPF 患者肺組織中 CAV1 表達明顯降低[12],這與本研究生物信息學分析結果一致。原因是正常表達 CAV1 可以抑制轉化生長因子(transforming growth factor, TGF)-β 的過表達,CAV1 表達降低會減弱 TGF-β 的抑制作用,從而激活 TGF-β 信號通路,導致細胞外基質大量生成,導致肺纖維化的發生[13]。本研究中的肺纖維化小鼠模型進一步顯示了在博來霉素誘導的肺纖維化組織中,CAV1 mRNA 的表達水平明顯低于正常對照組;總體生存分析顯示,CAV1 高表達患者的生存時間更長。CAV1 是纖維素的主要成分,調節細胞信號傳導和內吞作用,過表達 CAV1 可以減少中性粒細胞和單核/巨噬細胞的浸潤,防止博來霉素誘導的肺纖維化,這可能與其在炎癥活動中的關鍵調節作用有關[14-15]。本研究動物實驗表明,在肺纖維化小鼠中進行跑步機運動時,CAV1 mRNA 的表達增加。相關研究表明,在博來霉素誘導的肺纖維化模型中,跑步機運動可改善肺部和全身炎癥[14],這可能與本研究中跑步機增加 CAV1 有關。
CDKN2A 被稱為周期依賴性激酶抑制劑基因,屬于周期依賴性激酶抑制劑家族,是重要的腫瘤抑制基因,IPF 患者外周血 CDKN2A mRNA 表達水平較對照組降低,可能與 P53 信號通路激活有關[16]。在本實驗中,肺纖維化小鼠肺組織中 CDKN2A mRNA的表達水平低于正常對照組。FERDB 在線工具預測 CDKN2A 與鐵死亡相關,并將其歸類為鐵死亡驅動因子,CDKN2A 是否是 IPF 中鐵死亡的驅動因素尚未明確。因此,本研究結果表明 CDKN2A 影響肺纖維化的鐵死亡,其機制值得進一步研究。
NOS2 是氧化代謝的關鍵基因。既往研究報道,在肺纖維化病理生理條件下,存在異常高濃度的一氧化氮,這可能與誘導型一氧化氮合酶活性升高有關,過量的一氧化氮產生可能有助于纖維化的發展[17-18]。GDF15 是一種內分泌激素,是 TGF-β 超家族的成員,可促進腫瘤細胞的鐵死亡;GDF15 是一種上皮來源的分泌蛋白,可能是一種有用的上皮應激生物標志物,因為其表達增加與 IPF 患者預后較差相關[19],這與本研究當前生物信息學分析的結果一致。HNF4A 是一種核轉錄因子,以同型二聚體的形式與 DNA 結合,主要與肝臟纖維化的發生有關,敲除 HNF4A 可緩解肝纖維化[20],然而沒有關于 HNF4A 與肺纖維化相關性的研究報道。同樣,本研究生物信息學分析的動物實驗和生存分析表明,HNF4A 與肺纖維化無關聯。本研究生物信息學分析數據表明,IPF 組織中 NOS2 和 CAV1 下調,CDKN2A 和 GDF15 上調;但在博來霉素誘導的小鼠 IPF 組織中,NOS2 上調,CDKN2A 下調。實驗結果與生物信息學分析結果的不一致可能是由于以下原因:① 用于生物信息學分析的數據來自 IPF 患者,而實驗數據來自博來霉素誘導的小鼠,雖然人和小鼠具有很高的基因同源性,但由于物種不同,結果可能存在一定的偏差;② 基因擴增具有很大的時間和空間特異性,從而導致生物信息學分析與實驗結果不一致。
本研究有以下局限性:首先,從 IPF 患者肺組織和正常肺組織中獲取生物信息學結果,由于實驗條件和醫院規模的限制,臨床組織樣本收集不足進行驗證。此外,動物模型中不同鐵死亡基因表達水平的驗證,缺乏樞紐基因在肺纖維化細胞模型中的潛在研究。
綜上所述,通過生物信息學分析,本研究確定了 20 個肺纖維化中與鐵死亡相關的潛在基因。CAV1、NOS2、GDF15、CDKN2A 可能通過調控鐵死亡影響肺纖維化的發生。肺纖維化誘導后的有氧運動訓練可減輕肺鐵死亡,可能有助于肺纖維化的治療。
利益沖突:所有作者聲明不存在利益沖突。