引用本文: 張悅, 湯煒, 田衛東, 于湄. 真皮脂肪調控毛發生長的研究進展. 中國修復重建外科雜志, 2024, 38(5): 626-632. doi: 10.7507/1002-1892.202402092 復制
版權信息: ?四川大學華西醫院華西期刊社《中國修復重建外科雜志》版權所有,未經授權不得轉載、改編
毛囊作為皮膚的外胚層附屬器官,含有獨特的干細胞群,能夠不斷發生周期性變化以再生新毛發[1]。毛囊的生理活動由一系列高度協調的上皮-間充質相互作用所調控,其周圍的真皮組織中富含多種類型細胞,包括脂肪細胞、成纖維細胞、免疫細胞等,它們共同分泌多種生長因子、信號分子等物質,構成了毛囊周圍復雜的微環境,影響毛囊的生長發育和毛發周期變化[2]。真皮白色脂肪組織(white adipose tissue,WAT)與毛囊解剖結構相鄰,其變化直接影響正常毛發周期[3]。本文旨在闡述真皮脂肪的變化及其與毛發周期轉變之間的關系,為臨床治療脫發提供新的研究思路。
1 真皮WAT和毛發周期之間的動態同步變化
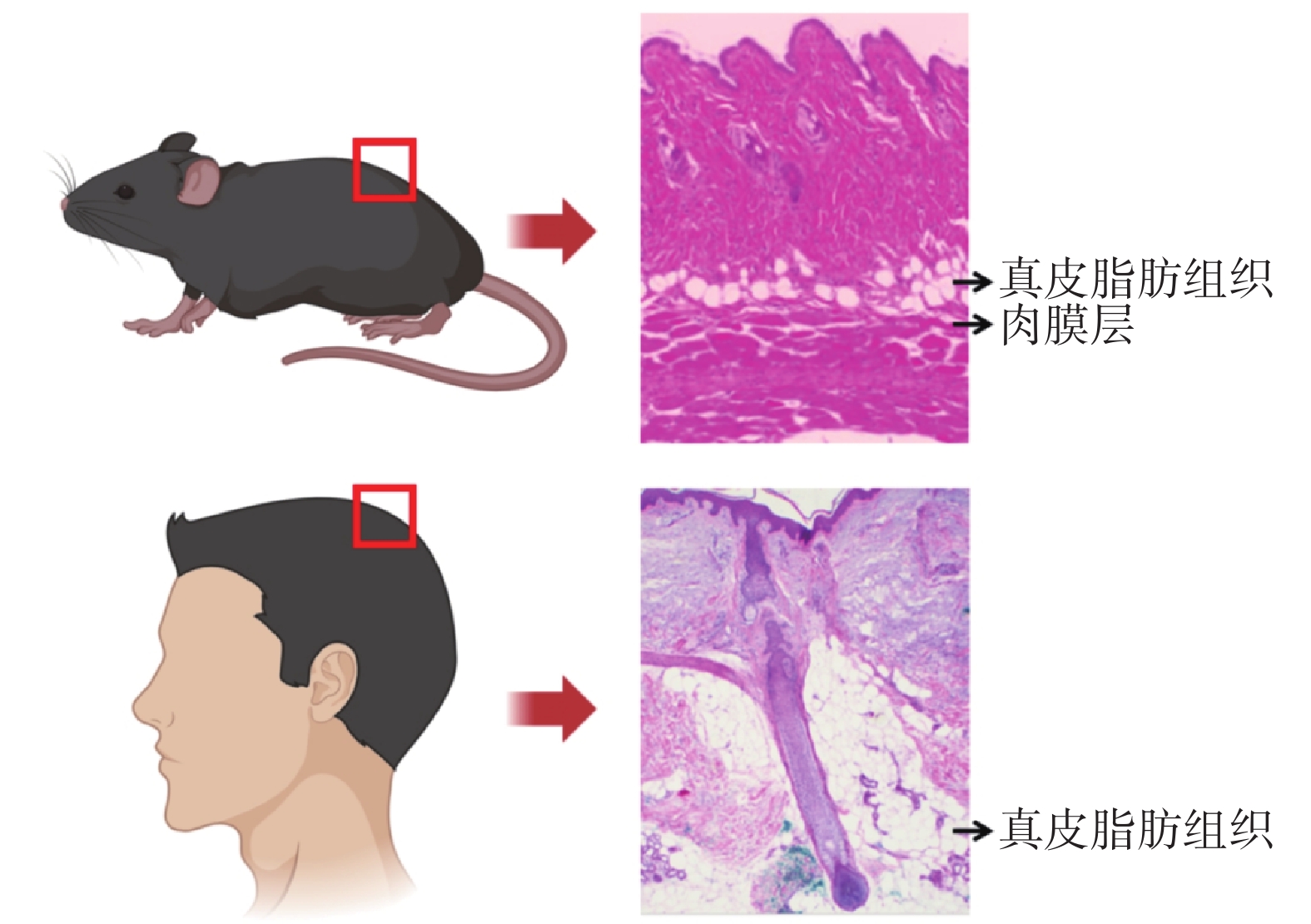
WAT廣泛分布于全身,它們既可以儲存營養來源的脂質,又可以根據需要進行動員,從而參與機體各項生理活動[4-5]。與體內其他主要WAT相似,皮膚WAT執行多種代謝功能(也稱為“系統功能”)及非代謝功能[6]。在人們早期認知中,皮膚相關的WAT統稱為“皮下脂肪”。然而,進一步研究發現,嚙齒動物網狀真皮下方存在兩層組織學和解剖學上各不相同的脂肪組織[7-10],二者通過中間的橫紋肌層(或稱“肉膜層”)分隔,肉膜層上方與表皮層相連的脂肪稱為“真皮WAT”,肉膜層下方的脂肪稱為“皮下脂肪”。人的皮膚和小鼠皮膚解剖結構不同,皮下無明顯肉膜層加以區分真皮脂肪,只能根據其解剖結構,將緊緊包繞毛囊并形成“真皮錐”結構的脂肪組織稱為“真皮WAT”,但缺乏明顯的形態學分隔與皮下WAT分界[11]。見圖1。
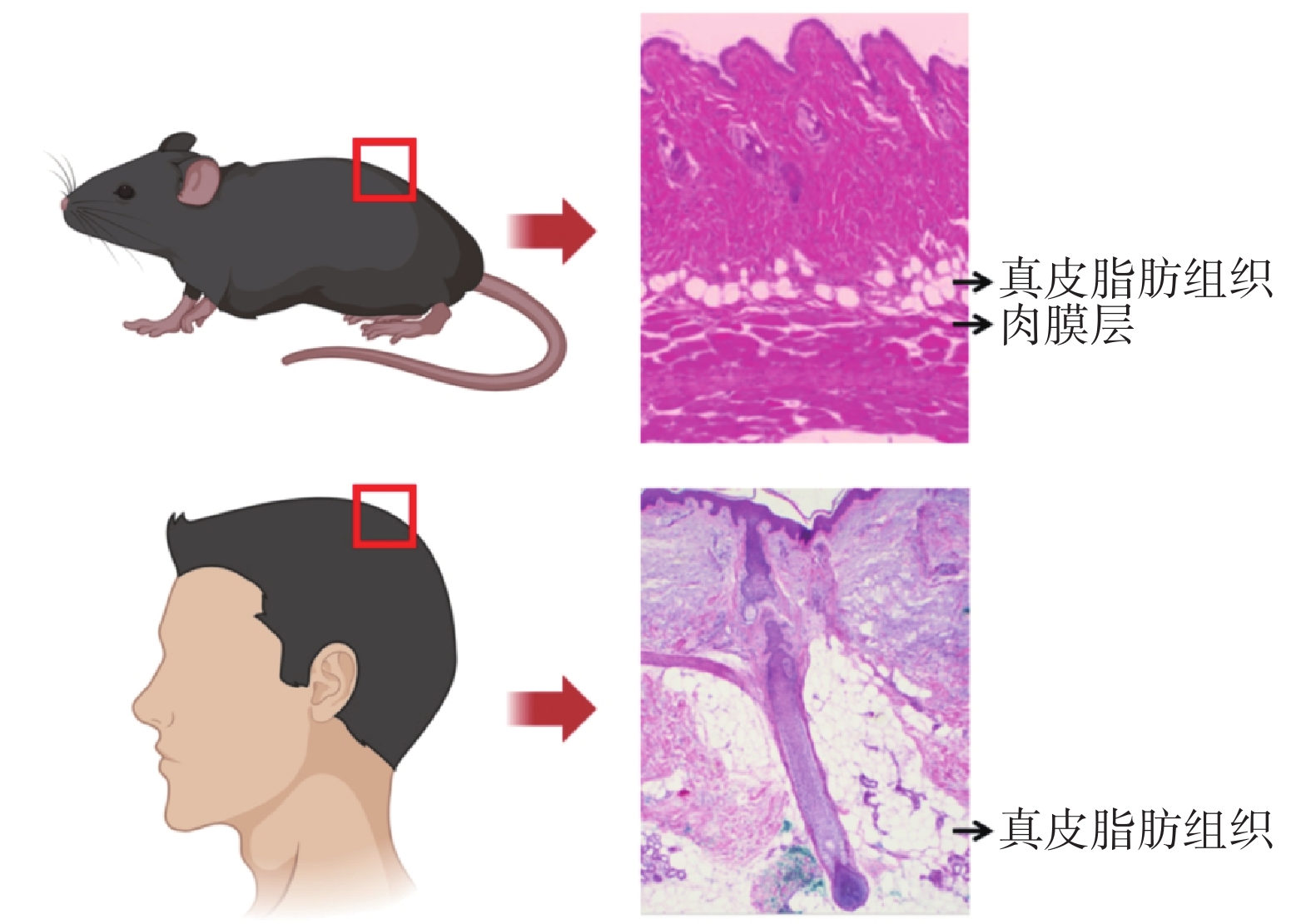 圖1
小鼠和人的真皮WAT結構差異(改編自參考文獻[12])
Figure1.
Structural differences of dermal WAT between mice and humans (adapted from reference [12])
圖1
小鼠和人的真皮WAT結構差異(改編自參考文獻[12])
Figure1.
Structural differences of dermal WAT between mice and humans (adapted from reference [12])
真皮WAT中存在成熟脂肪細胞、脂肪前體細胞、內皮細胞等多種細胞,并且不斷進行著脂肪生成和脂肪分解等代謝活動。脂肪前體細胞及成熟脂肪細胞的增殖代謝與毛囊周期變化之間存在緊密的時空相關性[12]。毛囊是皮膚的外胚層附屬器官,在毛發周期中反復再生新毛發。毛發周期包括生長期(生長階段)、退行期(退化階段)和靜止期(休息階段)。在靜止期晚期至生長期早期過渡期間,位于毛囊膨起部位的毛囊干細胞被位于毛囊基部的真皮乳頭細胞激活,進一步增殖、分化并向下遷移成為毛囊外根鞘細胞,而包繞真皮乳頭細胞的毛發基質細胞最終分化為毛干和內根鞘細胞,從而使毛囊逐漸向下伸長;毛囊退行期時,毛囊中的毛發基質細胞、內外根鞘細胞開始發生細胞凋亡,毛囊逐漸縮短,毛乳頭更靠近表皮層[13-14]。
緊鄰毛囊結構的真皮WAT與毛發周期循環同步發生變化。小鼠的皮下脂肪組織庫在出生后形成,而與毛發周期時空行為密切相關的只有真皮WAT,這也是真皮脂肪的標志性特征之一[15-17]。針對小鼠毛發周期的皮膚組織形態結構研究表明,真皮WAT與毛發周期同步變化體現在以下方面[12,18-21]:① 生長期:毛囊通過上皮細胞增殖向下伸長,真皮WAT層顯著增厚,充滿脂質的真皮成熟脂肪細胞圍繞生長中的毛囊;② 退行期:毛囊內發生細胞凋亡而向上縮短,真皮WAT厚度減小;③ 靜止期:在此期間毛囊較小且不活躍,真皮成熟脂肪細胞層減少至一層薄薄的成熟脂肪細胞。
有趣的是,除了皮膚解剖結構上的差異,人和嚙齒動物的毛發周期也大不相同。嚙齒動物毛囊的毛發周期在局部相鄰區域同時進行,這種現象造成了“馬賽克”樣皮膚毛發外觀的產生[22-24]。而人的毛發周期是單根毛囊獨立進行的,相鄰毛囊的毛發周期完全不同[25]。雖然毛發周期在不同種屬中表現形式不同,但真皮脂肪形態隨著毛發周期發生變化,即真皮WAT和毛發周期之間的動態同步在不同種屬中是相同的。
2 毛發周期中的脂肪前體細胞和成熟脂肪細胞
脂肪組織體積擴張表現為成熟脂肪細胞數量增多和體積增大,同時基質血管組分中的細胞組分也相應改變[4]。與毛發周期密切相關的真皮WAT的體積擴張,來自于真皮脂肪前體細胞增殖分化和真皮成熟脂肪細胞肥大的共同作用[26]。毛發生長期早期,真皮WAT體積擴張主要表現為成熟脂肪細胞數量增加[26]。驅動這一過程的是真皮脂肪前體細胞,包括脂肪干細胞(adipose-derived stem cells,ADSCs)及前成脂細胞。ADSCs屬于MSCs,主要存在于脂肪組織的基質血管組分,可以經誘導成為前成脂細胞[27-28]。前成脂細胞與ADSCs最大不同在于,ADSCs具有MSCs的多向分化潛能,但前成脂細胞是特定脂肪譜系的定向細胞群[29-30],只能增殖和分化為成熟脂肪細胞,并且具有成熟脂肪細胞的部分特征[31-33]。生長期早期脂肪前體細胞的數量為靜止期的4倍,在生長期晚期又恢復至靜止期水平[12]。
在毛發生長期中晚期,單個成熟脂肪細胞的體積增大是真皮WAT增厚的關鍵因素[34],成熟脂肪細胞數量及真皮WAT厚度均達峰值;當毛囊從生長期進入退行期時,真皮WAT的厚度降至其最大厚度的一半。這種真皮WAT厚度的變化是成熟脂肪細胞數量減少以及體積縮小共同作用的結果[15]。退行期中晚期成熟脂肪細胞內脂肪分解,體積減小[20,34];一部分成熟脂肪細胞還可以轉分化為成骨細胞、成纖維細胞等,另一部分能夠產生一群成熟脂肪細胞衍生的脂肪前體細胞[35],導致成熟脂肪細胞數量減少。
真皮成熟脂肪細胞并不是均質化細胞。首先,真皮成熟脂肪細胞群存在形成時間順序的異質性,新的毛發周期過程中所產生的成熟脂肪細胞與預先存在的成熟脂肪細胞并不是完全相同[28,35]。其次,不同成熟脂肪細胞對信號分子的敏感性存在差異。盡管靜止期時成熟脂肪細胞總數顯著下降,但部分成熟脂肪細胞在整個毛發周期中持續存在[12,16,26],這代表它們很難受到微環境刺激進而發生脂肪分解、轉分化等改變。第三,成熟脂肪細胞的異質性存在于空間水平上。成熟脂肪細胞沿毛囊長軸分布,靠近毛囊膨起部位毛囊干細胞的成熟脂肪細胞與位于毛囊基部毛乳頭細胞附近的成熟脂肪細胞空間位置不同,能夠響應不同的信號調節進而產生不同變化。
3 真皮脂肪對毛發生長的調控作用
3.1 真皮脂肪前體細胞
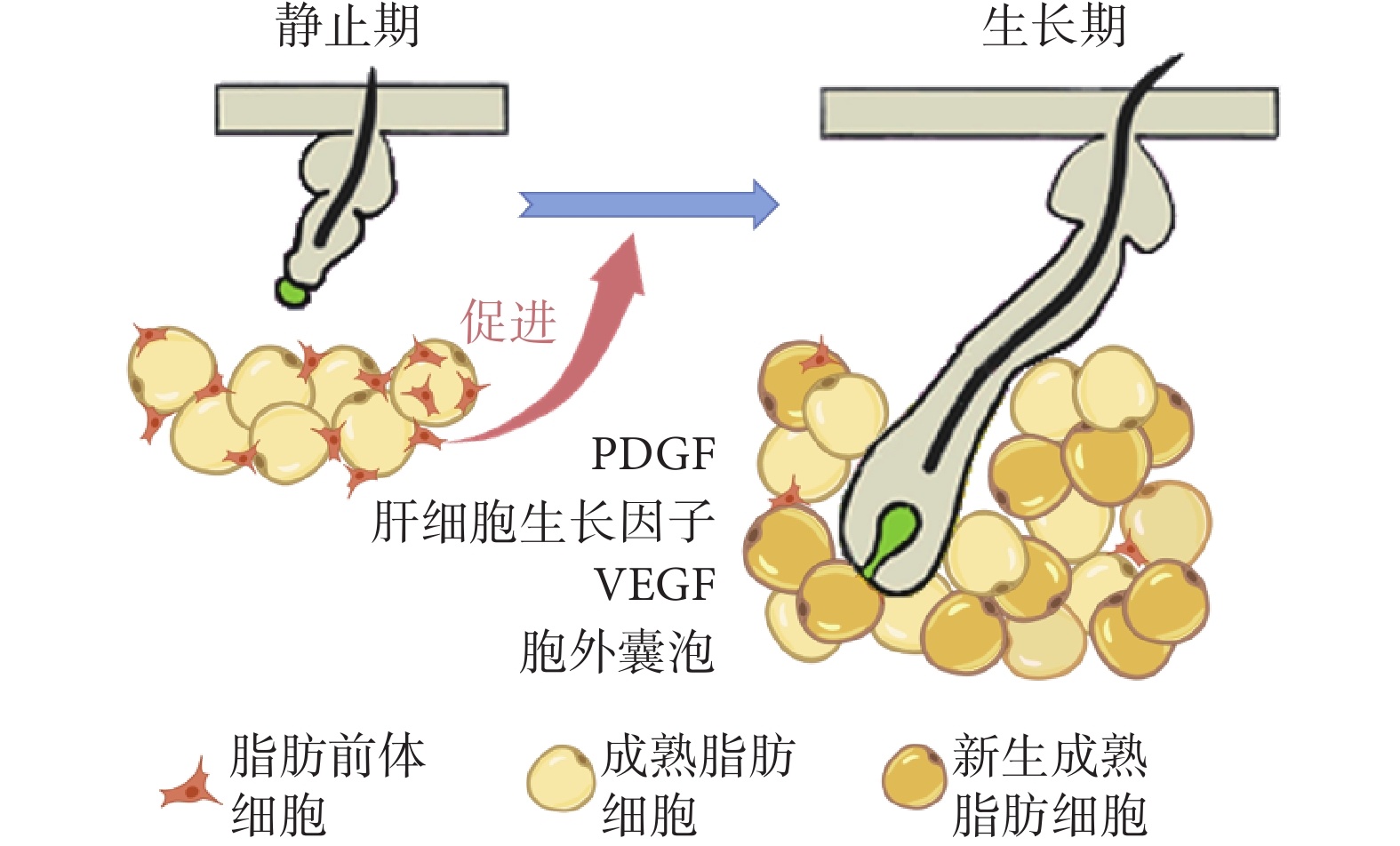
真皮脂肪前體細胞最明顯作用體現在毛發靜止期向生長期早期轉變過程中。真皮脂肪組織中具有增殖和分化能力的脂肪前體細胞數量增多時,毛囊加速進入生長期[12]。真皮脂肪前體細胞的增殖、分化驅動成熟脂肪細胞數量增加,真皮WAT體積擴張[12,27]。見圖2。
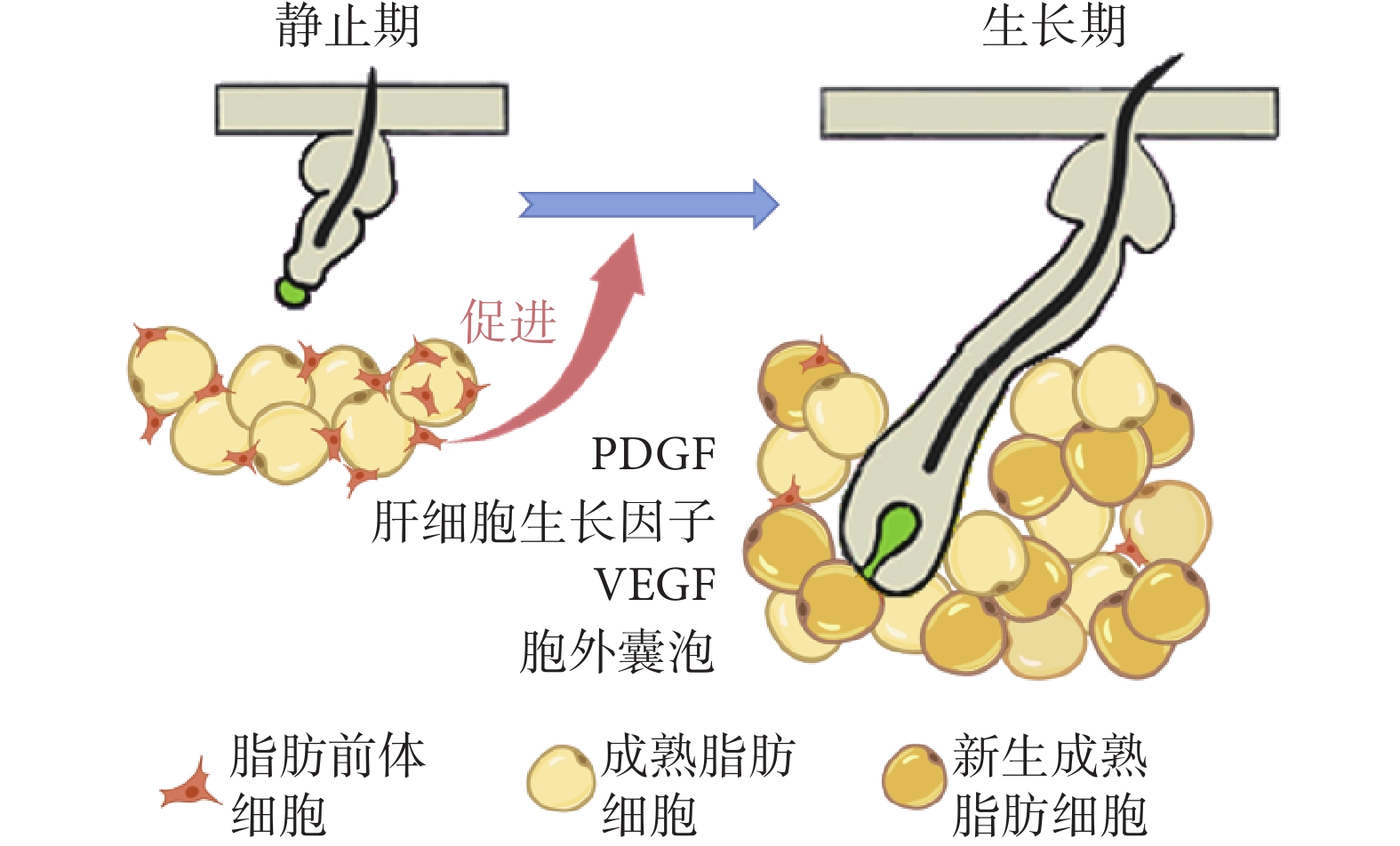 圖2
真皮脂肪前體細胞的增殖、分化加速毛發由靜止期向生長期轉變
Figure2.
The proliferation and differentiation of dermal adipose precursor cells accelerate the transition from telogen to anagen
圖2
真皮脂肪前體細胞的增殖、分化加速毛發由靜止期向生長期轉變
Figure2.
The proliferation and differentiation of dermal adipose precursor cells accelerate the transition from telogen to anagen
通過研究幾種脂肪前體細胞缺陷的突變小鼠可以明確真皮WAT與毛囊毛發周期的相關性。Festa等[12]的研究顯示,轉錄因子早期B-細胞因子1(early B-cell factor 1,Ebf1)基因敲除小鼠(Ebf1–/–)的皮膚在出生后表現出ADSCs缺乏、真皮脂肪組織變薄的特點,同時這些小鼠毛發周期停滯在退行期晚期或靜止期,不能重新進入生長期。他們將野生型小鼠的毛囊移植至Ebf1–/– 小鼠體內,發現其毛囊維持在靜止期狀態,無法進入生長期,毛發生長受到抑制,表明Ebf1–/– 小鼠的毛發生長缺陷可以歸因于脂肪前體細胞功能障礙。該研究還將來自野生型小鼠基質血管組分中富集的CD24陽性脂肪前體細胞移植至Ebf1–/– 小鼠皮膚中,發現小鼠背部皮膚黑色素沉積及毛發生長加快;但若僅移植脂肪組織中血管基質成分細胞,毛發周期變化不明顯。過氧化物酶體增殖物激活受體g(peroxisome proliferators-activated receptor g,PPARg)全缺失小鼠體內的活化PPARγ缺失,同時伴有脂肪前體細胞缺失,進一步導致毛囊發育不良、毛發周期紊亂和嚴重的毛囊周圍炎癥[36]。向小鼠腹腔注射PPARγ拮抗劑(如BADGE或GW9662)可造成皮膚內前成脂細胞缺失、成熟脂肪細胞形成過程阻斷,導致毛囊干細胞活化異常,毛發周期畸變[12]。
目前,脂肪前體細胞影響毛發周期的信號傳導機制還未完全解析,PDGF是其中重要一員。PDGF能夠介導毛囊干細胞的激活,對于維持毛發周期的正常進行必不可少。Festa等[12]將包被PDGF的0.1%牛血清白蛋白微球植入缺乏脂肪前體細胞的Ebf1–/– 小鼠皮膚內,發現可以激活小鼠毛囊的靜止期-生長期轉化;在毛發周期靜止期小鼠背部皮膚局部注射PDGF,10 d后小鼠背部皮膚出現黑色素沉積,毛發周期進入生長期。脂肪前體細胞能夠分泌較高水平的PDGF,進而加速毛囊的靜止期-生長期轉化。同時有研究發現[37-38],脂肪前體細胞能夠在毛囊和真皮脂肪組織共培養過程中分泌肝細胞生長因子,其受體c-Met在毛發基質細胞和真皮細胞中表達;脂肪前體細胞可以通過分泌肝細胞生長因子抑制毛乳頭中的分泌型卷曲相關蛋白1(secreted frizzled related protein 1,SFRP1)的表達,毛囊中經典Wnt信號通路被激活,進而促進毛囊干細胞的活化和毛囊角質形成細胞增殖,促進靜止期毛囊向生長期轉變。
脂肪前體細胞還能夠衍生VEGF,對毛囊周圍的血管形成至關重要。毛發生長靜止期時,脂肪前體細胞衍生的VEGF能夠增強內皮細胞中的VEGF信號傳導,顯著增強毛囊周圍血管形成,為毛囊干細胞提供具有豐富血液供應的微環境,以促進毛發周期向生長期轉變[11]。
脂肪前體細胞也能通過分泌胞外囊泡(exosomes,Exos)調節毛發生長。Wu等[39]將ADSCs來源的Exos(ADSC-Exos)、真皮細胞(dermal cells,DCs)和表皮細胞(epidermal cells,ECs)的混合物分別注射入小鼠背部皮膚,發現與DCs+ECs組相比,ADSC-Exos組背部更快出現毛發生長,毛發周期更早由靜止期進入生長期,且皮膚組織中PDGF及VEGF表達水平更高,而TGF-β1表達水平較低;此外,注射了ADSCs條件培養基的小鼠背部皮膚也更早出現毛發生長。
3.2 真皮成熟脂肪細胞
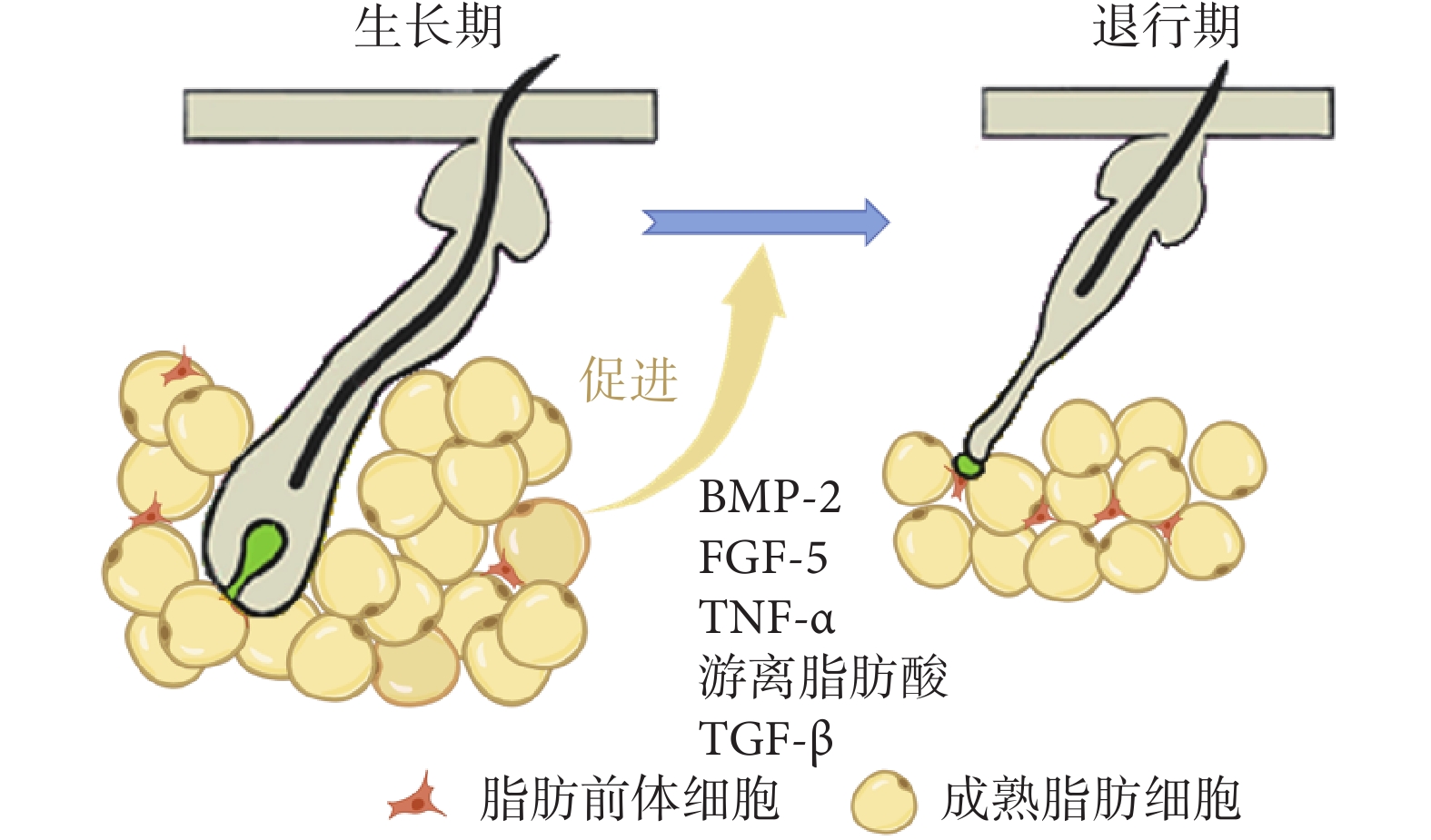
毛發生長期中晚期,單個成熟脂肪細胞體積增大是真皮WAT增厚的關鍵因素[27,40],成熟脂肪細胞數量及真皮脂肪組織層厚度均達到峰值。成熟脂肪細胞能夠起到縮短生長期、促進毛發周期向退行期轉變的作用。見圖3。
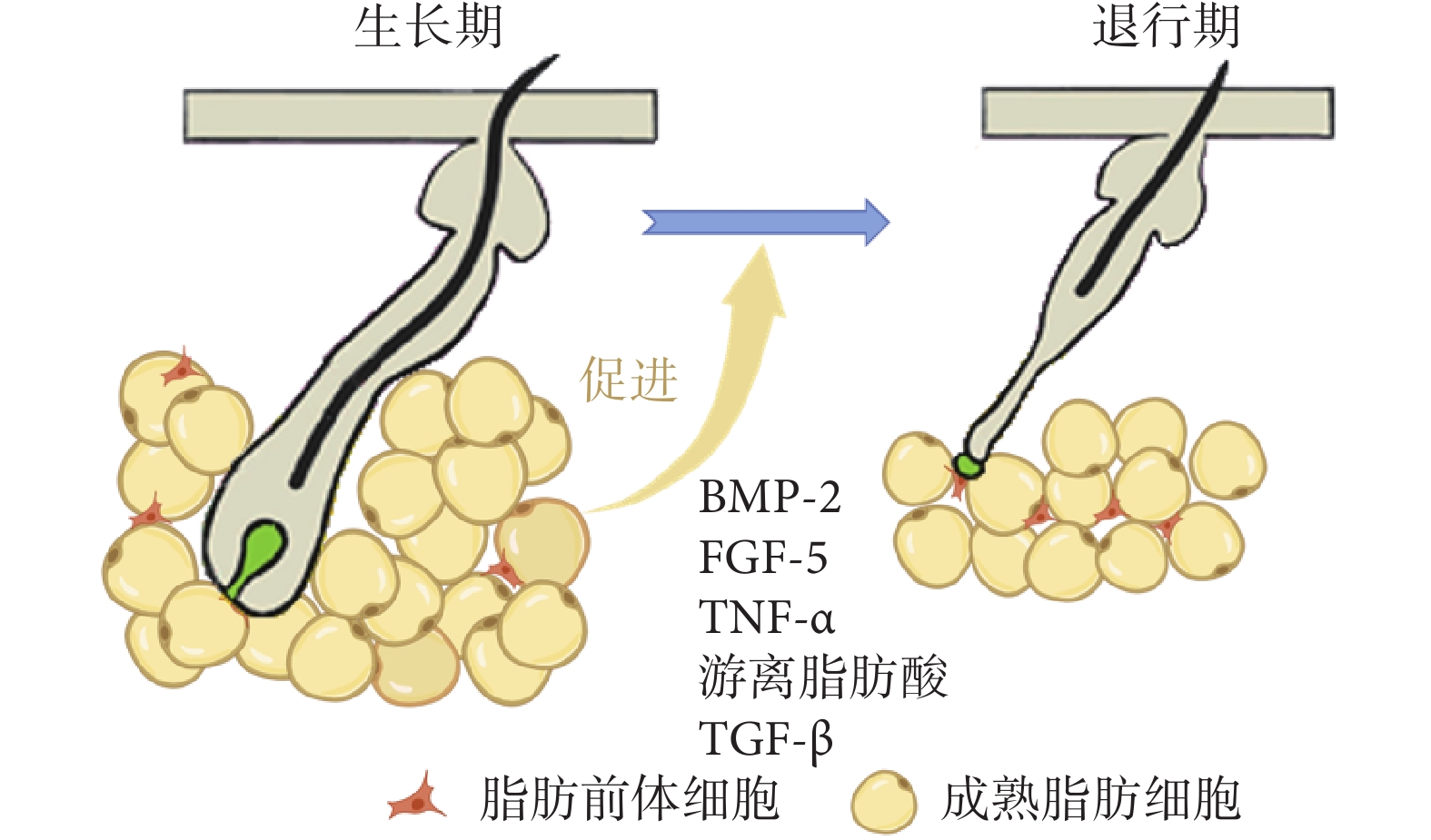 圖3
真皮成熟脂肪細胞在毛囊生長期-退行期轉變過程中的作用
Figure3.
The role of mature dermal adipocytes in the anagen-catagen transition of hair follicles
圖3
真皮成熟脂肪細胞在毛囊生長期-退行期轉變過程中的作用
Figure3.
The role of mature dermal adipocytes in the anagen-catagen transition of hair follicles
成熟脂肪細胞表達調節毛發周期的信號分子BMP-2,它在生長期晚期和靜止期早期達到峰值;在生長期晚期,BMP-2加速毛發周期向退行期轉變;而在靜止期早期,BMP-2抑制毛發重新進入生長期,保持毛囊的靜止狀態[41]。BMP信號轉導通過作用于pSMAD1/5/8相關靶基因來維持毛囊干細胞靜止狀態,縮短毛發生長期,加速其向退行期的轉變[42]。毛發生長期晚期,真皮WAT中的FGF水平顯著增加,其中FGF-5是毛囊生長的重要抑制因子,可誘導毛囊消退并促進其從生長期向退行期發生改變[43]。TNF-α信號轉導也會一定程度影響毛發周期變化,TNF-α的下游效應子與毛囊角蛋白17之間的分子串擾也可能通過調節細胞凋亡率來控制生長期的時長[44]。
脂肪分解產物游離脂肪酸是生長期晚期毛囊和脂肪細胞之間信號轉導的關鍵因素,它可以作為二級信使和配體,在細胞內和細胞間發揮作用。與正常小鼠6周齡時可觀察到毛發周期生長期-退行期轉變相比,高脂飲食喂養的小鼠皮膚內游離脂肪酸增多,能夠有效抑制毛囊干細胞中音猬因子(sonic hedgehog,SHH)信號傳導,促進毛囊細胞凋亡,毛囊更早出現生長期-退行期轉變[45]。皮膚中的游離脂肪酸還可能直接或間接促進毛囊或皮膚附屬器官中基因或蛋白質的表達,進而促進生長期毛囊轉變為退行期[46]。
毛囊生長期晚期,部分真皮成熟脂肪細胞開始去分化,這一過程能夠釋放高水平的TGF-β。TGF-β可以在毛囊底端提供病理微環境,使皮膚本身更容易受到炎癥影響,進而縮短生長期,加速毛發周期向退行期發生改變,并影響毛發周期靜止期,使之延長[47]。Foitzik等[48]將TGF-β1注射至小鼠背部皮膚中,成功誘導生長期毛囊的早期退行性變化,同時發現皮膚毛囊中增殖活躍的細胞數量減少、凋亡細胞數量增多。Tasaki等[49]發現肥胖小鼠毛囊的靜止期較長,肥大增生的成熟脂肪細胞能夠促進毛囊生長期-退行期變化。
同時,真皮成熟脂肪細胞還具有可塑性,它能夠通過轉分化變為成纖維細胞、成骨細胞等其他類型細胞[50]。真皮成熟脂肪細胞轉變為肌成纖維細胞,能夠引起小鼠皮膚纖維化[51];而小鼠皮膚的纖維化將進一步導致毛囊生長障礙,毛囊細胞發生凋亡[52]。
3.3 真皮脂肪前體細胞定向分化為成熟脂肪細胞
皮膚中的毛囊和真皮脂肪組織共同構成一個不斷同步變化,并涉及復雜信號相互作用的動態系統。脂肪前體細胞分化為成熟脂肪細胞的過程稱為“成脂分化”, 毛囊在不同毛發周期之間發生轉變及真皮成脂分化的過程共同受到多種信號通路的綜合調節,其中包括Wnt[53-58]、SHH[26,55,59-61]、BMP[62-67]、TGF-β[68-73]等信號通路。
經典Wnt信號通路在成脂分化及毛發周期中發揮重要作用。毛發生長期,上皮內經典Wnt信號通路被激活,抑制了其中的促脂肪轉錄因子CAAT增強子結合蛋白α(CAAT/enhancer-binding protein α,C/EBPα)和PPARγ的表達,進一步抑制了脂肪前體細胞的終末成脂分化過程[53]。在過度表達Wnt通路激活因子Wnt7a的小鼠皮膚中,脂肪前體細胞無法在生長期毛囊周圍增殖分化為成熟脂肪細胞[54]。Wnt7b不在靜止期表達,而是在出生后的第1個生長期開始表達,在出生后Wnt7b敲除小鼠中,毛囊干細胞的激活會受到抑制,毛發周期發生畸變[55]。同時,Wnt/β-catenin信號通路被認為是毛囊從靜止期轉變為生長期的中心信號通路,它的激活也能夠促進毛囊干細胞的活化、毛發周期向生長期轉變[56]。Wnt10b在Wnt/β-catenin信號通路中是不可或缺的。腺病毒介導的Wnt10b蛋白過表達可以誘導毛囊從靜息期過渡到生長期[57];Wnt10b的小干擾RNA介導的基因敲除小鼠體內毛囊無法進入生長期[58]。Wnt信號通路、負調節因子SFRP1的上調可以抑制毛囊干細胞增殖分化為毛囊外根鞘細胞[38]。
SHH信號通路的變化影響真皮成脂分化過程。生長期毛囊能夠通過分泌SHH配體來活化PPARγ,激活成脂分化過程[26]。當毛囊上皮中的SHH相關基因被敲除,或者當平滑蛋白特異性缺失的脂肪前體細胞對 SHH信號不敏感時,成脂分化被極大抑制;與之相反,SHH信號的皮膚特異性高表達會導致真皮脂肪層過度增厚,甚至達到正常厚度的2倍[26]。同時, SHH通路激活可誘導靜止干細胞的增殖,毛囊形態發生與 SHH信號密切相關[57]。SHH配體與其位于毛囊干細胞膜表面的受體Patched的結合能夠消除其對平滑蛋白的抑制作用,從而促進靶基因Gli1的轉錄以激活毛囊干細胞,促進毛發周期從靜止期向生長期的轉變[55,60-61]。
BMP信號通路是成脂分化的另一種重要調節因子。BMP信號通過激活轉錄調節因子鋅指蛋白423(zinc finger protein 423,ZFP423)促進脂肪前體細胞的終末分化[62];正常皮膚中的ZFP423缺失可導致真皮WAT明顯變薄和成熟脂肪細胞體積縮小[55]。用BMP-4處理C3H10T1/2多能細胞,能夠促進促脂肪轉錄因子C/EBPα和PPARγ的表達,使其轉化為成熟脂肪細胞[63]。同時,用BMP拮抗劑noggin處理來源于C3H10T1/2細胞的脂肪前體細胞,可抑制其進一步成脂分化過程[64]。BMP信號傳導是維持毛囊干細胞靜止狀態的關鍵機制,進而能夠將毛發周期維持在靜止期[65]。 減少或抑制BMP信號傳導可以激活毛囊干細胞,小鼠中的骨成型蛋白受體1α丟失可導致靜止毛囊干細胞的異常激活[66-67]。
TGF-β通過抑制C/EBPα和PPARγ的表達來抑制脂肪前體細胞的分化[68-69]。MSCs中TGF-β受體2缺失會導致小鼠骨髓中的PPARγ表達及脂肪組織擴增顯著增加[70]。de Ferranti等[71]將TGF-β1與人BMSCs共培養7 d,發現細胞成脂分化減少。此外,TGF-β1對C/EBPα和C/EBPβ的抑制增加了PPARγ的絲氨酸磷酸化,從而抑制了人BMSCs的成脂分化[59]。TGF-β信號通路同時也會對毛發周期產生影響,TGF-β能夠激活毛囊干細胞中的Smad信號,在靜止期至生長期過渡期間促進毛囊干細胞的激活[72-73]。
4 結語
毛囊和真皮WAT存在時空相互作用,生長期毛囊能夠促進脂肪前體細胞增殖分化為成熟脂肪細胞,而靜止期毛囊可介導真皮成熟脂肪細胞的脂肪分解。同時越來越多研究證實,脂肪前體細胞可激活靜止期毛囊向生長期轉變,成熟脂肪細胞能夠保持毛發周期的靜止狀態、抑制新一輪毛發周期。脂肪組織能夠調節毛發生長,但目前的研究尚不足以充分闡明脂肪組織在調節毛發生長周期中的突出作用,隨著更多調控毛發周期及脂肪組織變化的微環境信號被進一步研究,今后對于各類皮膚缺陷、毛發生長缺陷將出現更多更有效的臨床治療手段。
利益沖突 在課題研究和文章撰寫過程中不存在利益沖突;經費支持沒有影響文章觀點及其報道
作者貢獻聲明 張悅:文章構思及撰寫;湯煒:文獻整理匯總;田衛東:文章審閱及資金支持;于湄:文章審閱及修改
毛囊作為皮膚的外胚層附屬器官,含有獨特的干細胞群,能夠不斷發生周期性變化以再生新毛發[1]。毛囊的生理活動由一系列高度協調的上皮-間充質相互作用所調控,其周圍的真皮組織中富含多種類型細胞,包括脂肪細胞、成纖維細胞、免疫細胞等,它們共同分泌多種生長因子、信號分子等物質,構成了毛囊周圍復雜的微環境,影響毛囊的生長發育和毛發周期變化[2]。真皮白色脂肪組織(white adipose tissue,WAT)與毛囊解剖結構相鄰,其變化直接影響正常毛發周期[3]。本文旨在闡述真皮脂肪的變化及其與毛發周期轉變之間的關系,為臨床治療脫發提供新的研究思路。
1 真皮WAT和毛發周期之間的動態同步變化
WAT廣泛分布于全身,它們既可以儲存營養來源的脂質,又可以根據需要進行動員,從而參與機體各項生理活動[4-5]。與體內其他主要WAT相似,皮膚WAT執行多種代謝功能(也稱為“系統功能”)及非代謝功能[6]。在人們早期認知中,皮膚相關的WAT統稱為“皮下脂肪”。然而,進一步研究發現,嚙齒動物網狀真皮下方存在兩層組織學和解剖學上各不相同的脂肪組織[7-10],二者通過中間的橫紋肌層(或稱“肉膜層”)分隔,肉膜層上方與表皮層相連的脂肪稱為“真皮WAT”,肉膜層下方的脂肪稱為“皮下脂肪”。人的皮膚和小鼠皮膚解剖結構不同,皮下無明顯肉膜層加以區分真皮脂肪,只能根據其解剖結構,將緊緊包繞毛囊并形成“真皮錐”結構的脂肪組織稱為“真皮WAT”,但缺乏明顯的形態學分隔與皮下WAT分界[11]。見圖1。
圖1
小鼠和人的真皮WAT結構差異(改編自參考文獻[12])
Figure1.
Structural differences of dermal WAT between mice and humans (adapted from reference [12])
真皮WAT中存在成熟脂肪細胞、脂肪前體細胞、內皮細胞等多種細胞,并且不斷進行著脂肪生成和脂肪分解等代謝活動。脂肪前體細胞及成熟脂肪細胞的增殖代謝與毛囊周期變化之間存在緊密的時空相關性[12]。毛囊是皮膚的外胚層附屬器官,在毛發周期中反復再生新毛發。毛發周期包括生長期(生長階段)、退行期(退化階段)和靜止期(休息階段)。在靜止期晚期至生長期早期過渡期間,位于毛囊膨起部位的毛囊干細胞被位于毛囊基部的真皮乳頭細胞激活,進一步增殖、分化并向下遷移成為毛囊外根鞘細胞,而包繞真皮乳頭細胞的毛發基質細胞最終分化為毛干和內根鞘細胞,從而使毛囊逐漸向下伸長;毛囊退行期時,毛囊中的毛發基質細胞、內外根鞘細胞開始發生細胞凋亡,毛囊逐漸縮短,毛乳頭更靠近表皮層[13-14]。
緊鄰毛囊結構的真皮WAT與毛發周期循環同步發生變化。小鼠的皮下脂肪組織庫在出生后形成,而與毛發周期時空行為密切相關的只有真皮WAT,這也是真皮脂肪的標志性特征之一[15-17]。針對小鼠毛發周期的皮膚組織形態結構研究表明,真皮WAT與毛發周期同步變化體現在以下方面[12,18-21]:① 生長期:毛囊通過上皮細胞增殖向下伸長,真皮WAT層顯著增厚,充滿脂質的真皮成熟脂肪細胞圍繞生長中的毛囊;② 退行期:毛囊內發生細胞凋亡而向上縮短,真皮WAT厚度減小;③ 靜止期:在此期間毛囊較小且不活躍,真皮成熟脂肪細胞層減少至一層薄薄的成熟脂肪細胞。
有趣的是,除了皮膚解剖結構上的差異,人和嚙齒動物的毛發周期也大不相同。嚙齒動物毛囊的毛發周期在局部相鄰區域同時進行,這種現象造成了“馬賽克”樣皮膚毛發外觀的產生[22-24]。而人的毛發周期是單根毛囊獨立進行的,相鄰毛囊的毛發周期完全不同[25]。雖然毛發周期在不同種屬中表現形式不同,但真皮脂肪形態隨著毛發周期發生變化,即真皮WAT和毛發周期之間的動態同步在不同種屬中是相同的。
2 毛發周期中的脂肪前體細胞和成熟脂肪細胞
脂肪組織體積擴張表現為成熟脂肪細胞數量增多和體積增大,同時基質血管組分中的細胞組分也相應改變[4]。與毛發周期密切相關的真皮WAT的體積擴張,來自于真皮脂肪前體細胞增殖分化和真皮成熟脂肪細胞肥大的共同作用[26]。毛發生長期早期,真皮WAT體積擴張主要表現為成熟脂肪細胞數量增加[26]。驅動這一過程的是真皮脂肪前體細胞,包括脂肪干細胞(adipose-derived stem cells,ADSCs)及前成脂細胞。ADSCs屬于MSCs,主要存在于脂肪組織的基質血管組分,可以經誘導成為前成脂細胞[27-28]。前成脂細胞與ADSCs最大不同在于,ADSCs具有MSCs的多向分化潛能,但前成脂細胞是特定脂肪譜系的定向細胞群[29-30],只能增殖和分化為成熟脂肪細胞,并且具有成熟脂肪細胞的部分特征[31-33]。生長期早期脂肪前體細胞的數量為靜止期的4倍,在生長期晚期又恢復至靜止期水平[12]。
在毛發生長期中晚期,單個成熟脂肪細胞的體積增大是真皮WAT增厚的關鍵因素[34],成熟脂肪細胞數量及真皮WAT厚度均達峰值;當毛囊從生長期進入退行期時,真皮WAT的厚度降至其最大厚度的一半。這種真皮WAT厚度的變化是成熟脂肪細胞數量減少以及體積縮小共同作用的結果[15]。退行期中晚期成熟脂肪細胞內脂肪分解,體積減小[20,34];一部分成熟脂肪細胞還可以轉分化為成骨細胞、成纖維細胞等,另一部分能夠產生一群成熟脂肪細胞衍生的脂肪前體細胞[35],導致成熟脂肪細胞數量減少。
真皮成熟脂肪細胞并不是均質化細胞。首先,真皮成熟脂肪細胞群存在形成時間順序的異質性,新的毛發周期過程中所產生的成熟脂肪細胞與預先存在的成熟脂肪細胞并不是完全相同[28,35]。其次,不同成熟脂肪細胞對信號分子的敏感性存在差異。盡管靜止期時成熟脂肪細胞總數顯著下降,但部分成熟脂肪細胞在整個毛發周期中持續存在[12,16,26],這代表它們很難受到微環境刺激進而發生脂肪分解、轉分化等改變。第三,成熟脂肪細胞的異質性存在于空間水平上。成熟脂肪細胞沿毛囊長軸分布,靠近毛囊膨起部位毛囊干細胞的成熟脂肪細胞與位于毛囊基部毛乳頭細胞附近的成熟脂肪細胞空間位置不同,能夠響應不同的信號調節進而產生不同變化。
3 真皮脂肪對毛發生長的調控作用
3.1 真皮脂肪前體細胞
真皮脂肪前體細胞最明顯作用體現在毛發靜止期向生長期早期轉變過程中。真皮脂肪組織中具有增殖和分化能力的脂肪前體細胞數量增多時,毛囊加速進入生長期[12]。真皮脂肪前體細胞的增殖、分化驅動成熟脂肪細胞數量增加,真皮WAT體積擴張[12,27]。見圖2。
圖2
真皮脂肪前體細胞的增殖、分化加速毛發由靜止期向生長期轉變
Figure2.
The proliferation and differentiation of dermal adipose precursor cells accelerate the transition from telogen to anagen
通過研究幾種脂肪前體細胞缺陷的突變小鼠可以明確真皮WAT與毛囊毛發周期的相關性。Festa等[12]的研究顯示,轉錄因子早期B-細胞因子1(early B-cell factor 1,Ebf1)基因敲除小鼠(Ebf1–/–)的皮膚在出生后表現出ADSCs缺乏、真皮脂肪組織變薄的特點,同時這些小鼠毛發周期停滯在退行期晚期或靜止期,不能重新進入生長期。他們將野生型小鼠的毛囊移植至Ebf1–/– 小鼠體內,發現其毛囊維持在靜止期狀態,無法進入生長期,毛發生長受到抑制,表明Ebf1–/– 小鼠的毛發生長缺陷可以歸因于脂肪前體細胞功能障礙。該研究還將來自野生型小鼠基質血管組分中富集的CD24陽性脂肪前體細胞移植至Ebf1–/– 小鼠皮膚中,發現小鼠背部皮膚黑色素沉積及毛發生長加快;但若僅移植脂肪組織中血管基質成分細胞,毛發周期變化不明顯。過氧化物酶體增殖物激活受體g(peroxisome proliferators-activated receptor g,PPARg)全缺失小鼠體內的活化PPARγ缺失,同時伴有脂肪前體細胞缺失,進一步導致毛囊發育不良、毛發周期紊亂和嚴重的毛囊周圍炎癥[36]。向小鼠腹腔注射PPARγ拮抗劑(如BADGE或GW9662)可造成皮膚內前成脂細胞缺失、成熟脂肪細胞形成過程阻斷,導致毛囊干細胞活化異常,毛發周期畸變[12]。
目前,脂肪前體細胞影響毛發周期的信號傳導機制還未完全解析,PDGF是其中重要一員。PDGF能夠介導毛囊干細胞的激活,對于維持毛發周期的正常進行必不可少。Festa等[12]將包被PDGF的0.1%牛血清白蛋白微球植入缺乏脂肪前體細胞的Ebf1–/– 小鼠皮膚內,發現可以激活小鼠毛囊的靜止期-生長期轉化;在毛發周期靜止期小鼠背部皮膚局部注射PDGF,10 d后小鼠背部皮膚出現黑色素沉積,毛發周期進入生長期。脂肪前體細胞能夠分泌較高水平的PDGF,進而加速毛囊的靜止期-生長期轉化。同時有研究發現[37-38],脂肪前體細胞能夠在毛囊和真皮脂肪組織共培養過程中分泌肝細胞生長因子,其受體c-Met在毛發基質細胞和真皮細胞中表達;脂肪前體細胞可以通過分泌肝細胞生長因子抑制毛乳頭中的分泌型卷曲相關蛋白1(secreted frizzled related protein 1,SFRP1)的表達,毛囊中經典Wnt信號通路被激活,進而促進毛囊干細胞的活化和毛囊角質形成細胞增殖,促進靜止期毛囊向生長期轉變。
脂肪前體細胞還能夠衍生VEGF,對毛囊周圍的血管形成至關重要。毛發生長靜止期時,脂肪前體細胞衍生的VEGF能夠增強內皮細胞中的VEGF信號傳導,顯著增強毛囊周圍血管形成,為毛囊干細胞提供具有豐富血液供應的微環境,以促進毛發周期向生長期轉變[11]。
脂肪前體細胞也能通過分泌胞外囊泡(exosomes,Exos)調節毛發生長。Wu等[39]將ADSCs來源的Exos(ADSC-Exos)、真皮細胞(dermal cells,DCs)和表皮細胞(epidermal cells,ECs)的混合物分別注射入小鼠背部皮膚,發現與DCs+ECs組相比,ADSC-Exos組背部更快出現毛發生長,毛發周期更早由靜止期進入生長期,且皮膚組織中PDGF及VEGF表達水平更高,而TGF-β1表達水平較低;此外,注射了ADSCs條件培養基的小鼠背部皮膚也更早出現毛發生長。
3.2 真皮成熟脂肪細胞
毛發生長期中晚期,單個成熟脂肪細胞體積增大是真皮WAT增厚的關鍵因素[27,40],成熟脂肪細胞數量及真皮脂肪組織層厚度均達到峰值。成熟脂肪細胞能夠起到縮短生長期、促進毛發周期向退行期轉變的作用。見圖3。
圖3
真皮成熟脂肪細胞在毛囊生長期-退行期轉變過程中的作用
Figure3.
The role of mature dermal adipocytes in the anagen-catagen transition of hair follicles
成熟脂肪細胞表達調節毛發周期的信號分子BMP-2,它在生長期晚期和靜止期早期達到峰值;在生長期晚期,BMP-2加速毛發周期向退行期轉變;而在靜止期早期,BMP-2抑制毛發重新進入生長期,保持毛囊的靜止狀態[41]。BMP信號轉導通過作用于pSMAD1/5/8相關靶基因來維持毛囊干細胞靜止狀態,縮短毛發生長期,加速其向退行期的轉變[42]。毛發生長期晚期,真皮WAT中的FGF水平顯著增加,其中FGF-5是毛囊生長的重要抑制因子,可誘導毛囊消退并促進其從生長期向退行期發生改變[43]。TNF-α信號轉導也會一定程度影響毛發周期變化,TNF-α的下游效應子與毛囊角蛋白17之間的分子串擾也可能通過調節細胞凋亡率來控制生長期的時長[44]。
脂肪分解產物游離脂肪酸是生長期晚期毛囊和脂肪細胞之間信號轉導的關鍵因素,它可以作為二級信使和配體,在細胞內和細胞間發揮作用。與正常小鼠6周齡時可觀察到毛發周期生長期-退行期轉變相比,高脂飲食喂養的小鼠皮膚內游離脂肪酸增多,能夠有效抑制毛囊干細胞中音猬因子(sonic hedgehog,SHH)信號傳導,促進毛囊細胞凋亡,毛囊更早出現生長期-退行期轉變[45]。皮膚中的游離脂肪酸還可能直接或間接促進毛囊或皮膚附屬器官中基因或蛋白質的表達,進而促進生長期毛囊轉變為退行期[46]。
毛囊生長期晚期,部分真皮成熟脂肪細胞開始去分化,這一過程能夠釋放高水平的TGF-β。TGF-β可以在毛囊底端提供病理微環境,使皮膚本身更容易受到炎癥影響,進而縮短生長期,加速毛發周期向退行期發生改變,并影響毛發周期靜止期,使之延長[47]。Foitzik等[48]將TGF-β1注射至小鼠背部皮膚中,成功誘導生長期毛囊的早期退行性變化,同時發現皮膚毛囊中增殖活躍的細胞數量減少、凋亡細胞數量增多。Tasaki等[49]發現肥胖小鼠毛囊的靜止期較長,肥大增生的成熟脂肪細胞能夠促進毛囊生長期-退行期變化。
同時,真皮成熟脂肪細胞還具有可塑性,它能夠通過轉分化變為成纖維細胞、成骨細胞等其他類型細胞[50]。真皮成熟脂肪細胞轉變為肌成纖維細胞,能夠引起小鼠皮膚纖維化[51];而小鼠皮膚的纖維化將進一步導致毛囊生長障礙,毛囊細胞發生凋亡[52]。
3.3 真皮脂肪前體細胞定向分化為成熟脂肪細胞
皮膚中的毛囊和真皮脂肪組織共同構成一個不斷同步變化,并涉及復雜信號相互作用的動態系統。脂肪前體細胞分化為成熟脂肪細胞的過程稱為“成脂分化”, 毛囊在不同毛發周期之間發生轉變及真皮成脂分化的過程共同受到多種信號通路的綜合調節,其中包括Wnt[53-58]、SHH[26,55,59-61]、BMP[62-67]、TGF-β[68-73]等信號通路。
經典Wnt信號通路在成脂分化及毛發周期中發揮重要作用。毛發生長期,上皮內經典Wnt信號通路被激活,抑制了其中的促脂肪轉錄因子CAAT增強子結合蛋白α(CAAT/enhancer-binding protein α,C/EBPα)和PPARγ的表達,進一步抑制了脂肪前體細胞的終末成脂分化過程[53]。在過度表達Wnt通路激活因子Wnt7a的小鼠皮膚中,脂肪前體細胞無法在生長期毛囊周圍增殖分化為成熟脂肪細胞[54]。Wnt7b不在靜止期表達,而是在出生后的第1個生長期開始表達,在出生后Wnt7b敲除小鼠中,毛囊干細胞的激活會受到抑制,毛發周期發生畸變[55]。同時,Wnt/β-catenin信號通路被認為是毛囊從靜止期轉變為生長期的中心信號通路,它的激活也能夠促進毛囊干細胞的活化、毛發周期向生長期轉變[56]。Wnt10b在Wnt/β-catenin信號通路中是不可或缺的。腺病毒介導的Wnt10b蛋白過表達可以誘導毛囊從靜息期過渡到生長期[57];Wnt10b的小干擾RNA介導的基因敲除小鼠體內毛囊無法進入生長期[58]。Wnt信號通路、負調節因子SFRP1的上調可以抑制毛囊干細胞增殖分化為毛囊外根鞘細胞[38]。
SHH信號通路的變化影響真皮成脂分化過程。生長期毛囊能夠通過分泌SHH配體來活化PPARγ,激活成脂分化過程[26]。當毛囊上皮中的SHH相關基因被敲除,或者當平滑蛋白特異性缺失的脂肪前體細胞對 SHH信號不敏感時,成脂分化被極大抑制;與之相反,SHH信號的皮膚特異性高表達會導致真皮脂肪層過度增厚,甚至達到正常厚度的2倍[26]。同時, SHH通路激活可誘導靜止干細胞的增殖,毛囊形態發生與 SHH信號密切相關[57]。SHH配體與其位于毛囊干細胞膜表面的受體Patched的結合能夠消除其對平滑蛋白的抑制作用,從而促進靶基因Gli1的轉錄以激活毛囊干細胞,促進毛發周期從靜止期向生長期的轉變[55,60-61]。
BMP信號通路是成脂分化的另一種重要調節因子。BMP信號通過激活轉錄調節因子鋅指蛋白423(zinc finger protein 423,ZFP423)促進脂肪前體細胞的終末分化[62];正常皮膚中的ZFP423缺失可導致真皮WAT明顯變薄和成熟脂肪細胞體積縮小[55]。用BMP-4處理C3H10T1/2多能細胞,能夠促進促脂肪轉錄因子C/EBPα和PPARγ的表達,使其轉化為成熟脂肪細胞[63]。同時,用BMP拮抗劑noggin處理來源于C3H10T1/2細胞的脂肪前體細胞,可抑制其進一步成脂分化過程[64]。BMP信號傳導是維持毛囊干細胞靜止狀態的關鍵機制,進而能夠將毛發周期維持在靜止期[65]。 減少或抑制BMP信號傳導可以激活毛囊干細胞,小鼠中的骨成型蛋白受體1α丟失可導致靜止毛囊干細胞的異常激活[66-67]。
TGF-β通過抑制C/EBPα和PPARγ的表達來抑制脂肪前體細胞的分化[68-69]。MSCs中TGF-β受體2缺失會導致小鼠骨髓中的PPARγ表達及脂肪組織擴增顯著增加[70]。de Ferranti等[71]將TGF-β1與人BMSCs共培養7 d,發現細胞成脂分化減少。此外,TGF-β1對C/EBPα和C/EBPβ的抑制增加了PPARγ的絲氨酸磷酸化,從而抑制了人BMSCs的成脂分化[59]。TGF-β信號通路同時也會對毛發周期產生影響,TGF-β能夠激活毛囊干細胞中的Smad信號,在靜止期至生長期過渡期間促進毛囊干細胞的激活[72-73]。
4 結語
毛囊和真皮WAT存在時空相互作用,生長期毛囊能夠促進脂肪前體細胞增殖分化為成熟脂肪細胞,而靜止期毛囊可介導真皮成熟脂肪細胞的脂肪分解。同時越來越多研究證實,脂肪前體細胞可激活靜止期毛囊向生長期轉變,成熟脂肪細胞能夠保持毛發周期的靜止狀態、抑制新一輪毛發周期。脂肪組織能夠調節毛發生長,但目前的研究尚不足以充分闡明脂肪組織在調節毛發生長周期中的突出作用,隨著更多調控毛發周期及脂肪組織變化的微環境信號被進一步研究,今后對于各類皮膚缺陷、毛發生長缺陷將出現更多更有效的臨床治療手段。
利益沖突 在課題研究和文章撰寫過程中不存在利益沖突;經費支持沒有影響文章觀點及其報道
作者貢獻聲明 張悅:文章構思及撰寫;湯煒:文獻整理匯總;田衛東:文章審閱及資金支持;于湄:文章審閱及修改